
ХТО КЕРУЄ СТРЕСОМ — ТОЙ КЕРУЄ ВРОЖАЄМ
03.03.2026

ВРОЖАЙ ПОЧИНАЄТЬСЯ З НАУКИ.
ПРИБУТОК — З ПРАВИЛЬНИХ РІШЕНЬ.
Перша частина статті присвячена формуванню нової фізіолого-агрономічної парадигми управління врожайністю через контроль енергетичного стану рослини. У ній обґрунтовується, що сучасне рослинництво працює в умовах хронічного абіотичного навантаження, і саме фізіологічний стрес, а не лише хвороби чи шкідники, є головною причиною нестабільності врожаю. Показано, що більшість втрат продуктивності пов’язані не з прямим пошкодженням тканин, а з внутрішнім енергетичним дефіцитом клітини, який виникає під дією посухи, температурних коливань, порушення мінерального живлення та інших факторів. Стрес розглядається не як окремий зовнішній вплив, а як стан порушення балансу між утворенням і використанням енергії, що призводить до зміни метаболічних і гормональних програм.

Фото 1. Стрес як особливий фізіологічний стан, який формується всередині рослини.
У центрі уваги — механізми клітинного «прийняття рішення» між ростом і захистом. Детально аналізується роль енергетичних сенсорів TOR і SnRK1 як ключових регуляторів, що визначають, чи спрямовуватимуться ресурси на синтез біомаси та формування врожаю, чи на підтримку виживання. Розкривається взаємодія цих сенсорів із гормональними сигналами (цитокінінами, абсцизовою кислотою, жасмонатами, саліциловою кислотою), редокс-станом клітини та механізмами автофагії. Показано, що перемикання між режимами TOR-домінантного росту та SnRK1/ABA-домінантного захисту безпосередньо впливає на баланс SOURCE–SINK, силу наливу та реалізацію генетичного потенціалу культури.
Окремо підкреслюється, що зниження врожайності часто є наслідком стратегічного перерозподілу енергії всередині рослини: обмежується активність фотосинтезу, знижується транспорт асимілятів, слабшає сила поглинаючих органів. Таким чином, втрата врожаю розглядається як фізіологічний наслідок енергетичного перемикання, а не лише як прямий результат дії стресового чинника. Перша частина формує концепцію «енергетичного порогу», нижче якого ріст гальмується, і закладає теоретичну основу для переходу від реактивного захисту рослин до керованого управління їхнім метаболічним станом з метою стабілізації та підвищення врожайності.
Вступ
Світове рослинництво вступає в нову фазу розвитку, у якій традиційні агротехнології вже не забезпечують гарантованої стабільності врожаю. Кліматичні аномалії, хронічний вплив абіотичних стресів, деградація ґрунтів і підвищення агресивності патогенних організмів формують принципово нові умови існування агроценозів. У такій реальності рослина протягом значної частини вегетації функціонує не в оптимальному режимі росту і накопичення біомаси, а в режимі адаптації та виживання. Внаслідок цього класичні системи живлення і захисту, які раніше забезпечували стабільний результат, дедалі частіше демонструють нестабільну ефективність або не дозволяють повністю реалізувати генетичний потенціал культури.
За узагальненими оцінками міжнародних наукових і аналітичних джерел, саме абіотичні стреси є головною причиною глобальних втрат урожаю, формуючи близько 60–70% загального недобору продукції, тоді як частка біотичних чинників — хвороб і шкідників — становить лише 30–40%. Це свідчить про те, що основні втрати врожайності виникають задовго до появи видимих симптомів ураження, на рівні порушення енергетичного балансу, гормональної регуляції та базових метаболічних процесів. Таким чином, ключовим обмежувальним фактором сучасного агровиробництва стають не окремі патогени, а сукупний фізіологічний тиск середовища, який системно знижує ефективність функціонування рослинного організму.
У сучасних технологіях вирощування сільськогосподарських культур основний акцент традиційно зосереджений на контролі біотичних чинників — патогенів, шкідників і бур’янів. Саме ці фактори мають чіткі зовнішні прояви, легко діагностуються і, відповідно, для них створено широкий спектр засобів захисту з прогнозованою ефективністю. Водночас абіотичні фактори — посуха, температурні коливання, гербіцидний стрес, дефіцит елементів живлення, порушення водного та іонного балансу — які фактично формують основну частку втрат урожайності, у більшості технологічних схем залишаються поза системним контролем. Це створює фундаментальний дисбаланс у практиці сучасного землеробства, коли контролюється другорядна частина ризиків, тоді як домінуючий компонент втрат залишається некерованим.
Головною причиною такої ситуації є відсутність цілісної теоретичної концепції імунітету рослин, яка б розглядала захисні реакції як єдину інтегровану систему, спрямовану не лише проти біотичних агентів, але й проти абіотичних стресових факторів. У класичному розумінні імунітет рослин тривалий час асоціювався виключно з розпізнаванням і нейтралізацією патогенів, тоді як реакції на абіотичні стреси розглядалися як окремі фізіологічні або метаболічні адаптації, а не як складова єдиної захисної системи. Проте на клітинному і молекулярному рівнях відповідь на біотичні й абіотичні стреси має спільну природу і базується на єдиних регуляторних механізмах, включаючи перебудову енергетичного обміну, зміну гормонального балансу, регуляцію редокс-гомеостазу та активацію захисних метаболічних програм.
Відсутність такої інтегрованої теорії безпосередньо вплинула на розвиток практичних технологій і стала причиною того, що більшість наявних препаратів створені за принципом боротьби з наслідками, а не управління фундаментальними механізмами стресостійкості. На сьогодні практично відсутні препарати з чітко визначеною таргетною, тобто молекулярно специфічною, дією, спрямованою на регуляцію ключових внутрішньоклітинних систем стресової відповіді, таких як енергетичні сенсори, редокс-регулятори, сигнальні каскади та механізми підтримання клітинного гомеостазу. У результаті більшість застосовуваних продуктів виконують допоміжну або відновлювальну функцію, але не забезпечують повноцінного керування стресовою фізіологією рослини.
Таким чином, сучасне рослинництво фактично перебуває на етапі переходу від емпіричного контролю окремих факторів до формування нової науково обґрунтованої системи управління стійкістю рослин, у центрі якої має знаходитися цілеспрямована регуляція фундаментальних механізмів клітинної адаптації. Саме розвиток цього напрямку відкриває можливості для створення принципово нових технологічних рішень, здатних не лише зменшувати наслідки стресу, але й підтримувати рослину в активному продуктивному стані навіть за несприятливих умов середовища.
Економічний масштаб цієї проблеми є надзвичайно значним. За оцінками FAO, щорічні глобальні втрати врожаю від абіотичних стресів перевищують 99 млрд доларів США. Водночас це стимулює стрімкий розвиток ринку антистресових технологій, який уже сьогодні є одним із найбільш динамічних сегментів агроіндустрії. Прогнозується, що його обсяг зросте з 4,5 млрд доларів у 2025 році до майже 7,8 млрд доларів у 2030 році, що відображає фундаментальну зміну підходів до управління продуктивністю рослин.
До основних абіотичних викликів належать дефіцит води і порушення водного режиму, температурні стреси — як перегрів, так і переохолодження, засолення ґрунтів, порушення іонного гомеостазу, деградація ґрунтового середовища, зміни кислотності, накопичення токсичних сполук і хронічний оксидативний стрес. Принципова особливість цих факторів полягає в їх постійному і кумулятивному характері дії. Вони часто не викликають негайних візуальних симптомів, але поступово дестабілізують фізіологічні процеси, формуючи прихований дефіцит продуктивності, який проявляється значно пізніше.
На клітинному рівні абіотичний стрес є системним дестабілізуючим фактором, що порушує фундаментальні механізми функціонування рослини. Його дія супроводжується зниженням ефективності фотосинтезу, пригніченням синтезу АТФ, накопиченням реактивних форм кисню, зміною гормонального балансу та ослабленням захисних бар’єрів клітини. У результаті відбувається перехід від метаболічного режиму, спрямованого на ріст і продуктивність, до режиму збереження життєздатності. Навіть за відсутності явних симптомів це обмежує реалізацію потенціалу врожайності.
Численні польові дослідження підтверджують, що сукупна дія абіотичних і біотичних стресів може знижувати врожайність основних польових культур на 20–60%, а за екстремальних умов — ще більше. Найбільш поширеними факторами є посуха, температурні коливання, гербіцидне навантаження, дефіцит елементів живлення та пошкодження кореневої системи.
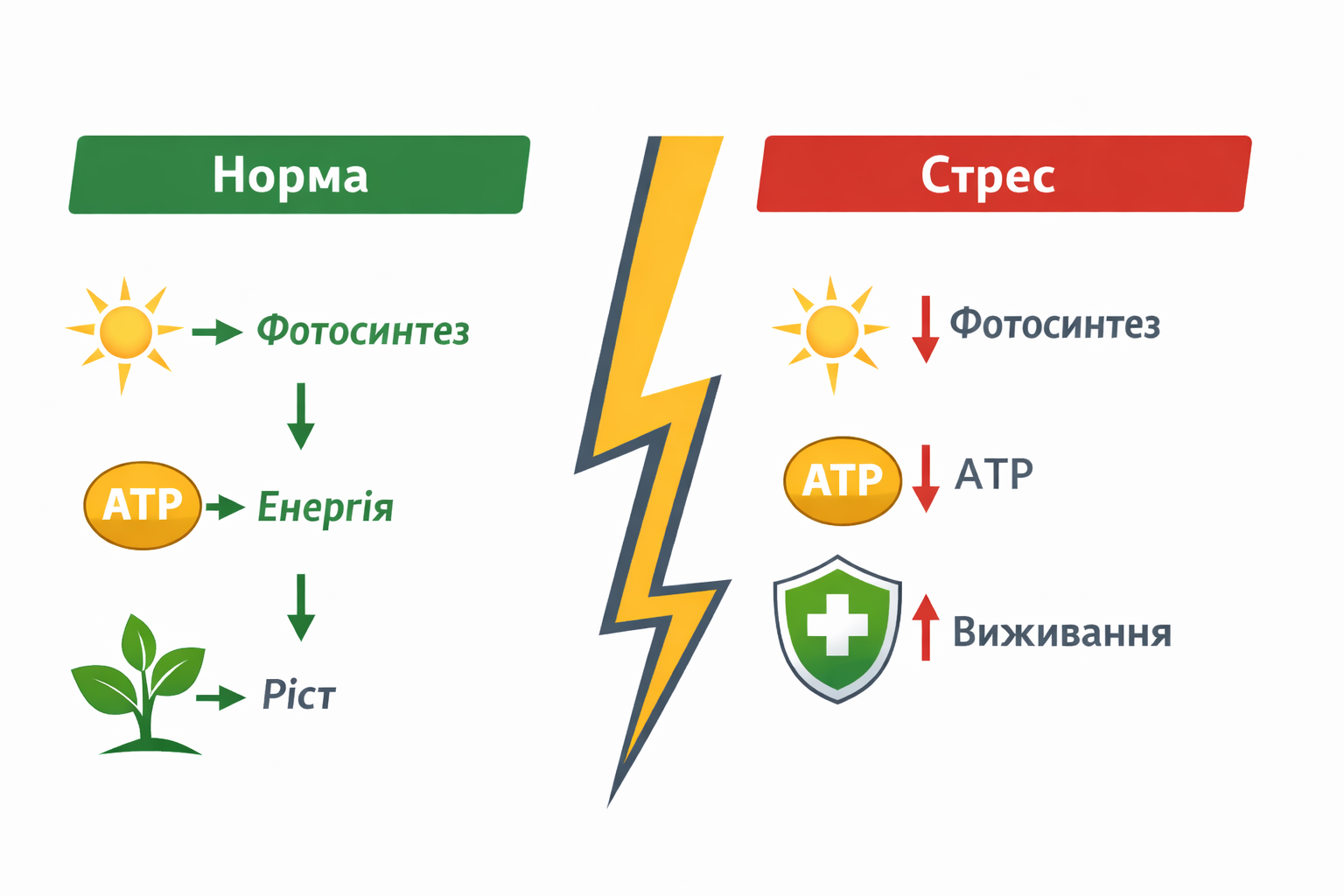
Фундаментальною причиною цих втрат є порушення енергетичного балансу рослини. У фізіологічному сенсі стрес можна розглядати як стан енергетичної кризи, при якому зменшується виробництво енергії та одночасно зростають витрати на підтримання захисних механізмів. У цих умовах рослина змушена перерозподіляти ресурси від процесів росту і формування врожаю до процесів адаптації і виживання, що безпосередньо визначає зниження продуктивності.
У зв’язку з цим сучасна технологія вирощування повинна виходити за межі традиційного забезпечення мінерального живлення. Її ключовим завданням стає управління фізіологічним і енергетичним станом рослини, зокрема підтримання ефективності фотосинтезу, стабілізація метаболізму і підвищення стійкості до стресових факторів. Це досягається через використання антистресових, імуномодулюючих і метаболічно активних продуктів, які дозволяють підтримувати функціональну стабільність рослин навіть у несприятливих умовах.
Важливо підкреслити, що стрес слід розглядати не лише як наслідок дії зовнішнього чинника, а як особливий фізіологічний стан, який формується всередині рослини. У цьому стані відбувається комплексна перебудова гормональних, сигнальних і метаболічних мереж, що визначає новий баланс між ростом і захистом. Такий підхід відображає сучасний парадигмальний зсув у розумінні природи стресу: він є не просто реакцією на зовнішній вплив, а інтегрованою програмою адаптації, яка визначає продуктивність рослини в умовах мінливого середовища. Саме тому управління стресом сьогодні стає одним із ключових інструментів підвищення врожайності, а ефективний контроль енергетичного і метаболічного стану рослини — центральним елементом сучасних інтенсивних агротехнологій.
Енергетична модель стресу та інтегрована модель імунітету рослин до патогенів і стресів «SA–JA/ET–ABA–TOR».
Енергетична модель стресу є фундаментальною концепцією сучасної фізіології рослин, яка розглядає стрес не лише як результат дії несприятливого фактора, а насамперед як наслідок порушення енергетичного балансу клітини. Згідно з цією моделлю, будь-який стресовий вплив — посуха, температурні екстремуми, засолення, дефіцит елементів живлення або ураження патогенами — призводить до ситуації, коли витрати енергії на підтримання життєво важливих процесів, захист і адаптацію перевищують можливості клітини щодо її синтезу. Таким чином, стрес по суті є станом енергетичного дефіциту, при якому порушується баланс між процесами генерації енергії (передусім фотосинтезом і диханням) та її споживанням. У нормальних умовах рослина підтримує стабільний енергетичний гомеостаз, що забезпечує ріст, синтез біомаси та формування врожаю. Проте за умов стресу відбувається різке зниження ефективності фотосинтетичного апарату, пошкодження мітохондріальних функцій і одночасне зростання енергетичних витрат на репарацію клітинних структур, детоксикацію активних форм кисню та активацію захисних механізмів. У результаті формується системний енергетичний дефіцит, який і є ключовою причиною пригнічення росту, втрати продуктивності та розвитку стресових симптомів. Саме тому енергетична модель стресу сьогодні розглядається як центральна парадигма, що об’єднує різні типи стресових реакцій у єдину універсальну систему, пояснюючи, що первинною причиною втрати продуктивності є не сам стресовий фактор, а викликане ним порушення енергетичного забезпечення клітини та рослини в цілому.
Мал. 2. Концептуальна схема стресу «Стрес як енергетичний дисбаланс».
Енергетична модель стресу вже частково розроблена, проте вона ще далека від завершення і містить фундаментальні прогалини, що відкривають потенціал для відкриттів. На сьогодні ми добре розуміємо окремі її компоненти, але головне питання залишається невирішеним: як саме клітина приймає рішення «жити, рости або виживати», виходячи з власного енергетичного стану.
Сучасна енергетична модель базується на кількох встановлених принципах. По-перше, АТФ є універсальною енергетичною валютою, відкриття хеміосмотичного механізму Пітера Мітчелла дозволило зрозуміти фундаментальні процеси її генерації. Відомо, що фотосинтез і мітохондрії виробляють АТФ, стресові умови знижують його синтез, а активні захисні механізми потребують енергії для підтримки життєдіяльності клітини.
По-друге, існують ключові енергетичні сенсори клітини. SnRK1 виступає сенсором дефіциту енергії, координуючи процеси виживання, катаболізму та зупинки росту, тоді як TOR реагує на надлишок енергії, стимулюючи ріст, синтез білка та накопичення біомаси. По-третє, стрес можна представити як зсув балансу між TOR і SnRK1, і цей принцип вже добре підтверджений експериментально.
Але головне питання залишається відкритим. Клітина не вимірює лише АТФ, вона оцінює загальний енергетичний стан системи, проте досі невідомо, який саме сигнал є критичним. Це може бути співвідношення ATP/ADP, ATP/AMP, NADH/NAD+, редокс стан, протонний градієнт, метаболічний потік або щось ще невідкрите. Найбільша прогалина сучасної науки полягає в тому, що ми не знаємо, як клітина інтегрує всі ці енергетичні сигнали в єдине рішення: рости, зупинитися, перейти в захист або вступити в програмовану смерть.
Найбільш перспективні напрями подальших відкриттів охоплюють кілька ключових процесів, які ще належить з’ясувати. По-перше, йдеться про ідентифікацію так званого «енергетичного вимикача клітини» — універсального порога енергетичного стану, нижче якого ріст зупиняється; наразі існування такого порога залишається гіпотетичним. По-друге, необхідно встановити первинний сенсор енергетичного дефіциту. Відомо, що SnRK1 функціонує як вторинний інтегратор сигналу, однак механізм його первинної активації досі не з’ясований. По-третє, практично нерозкритим залишається механізм перерозподілу енергії між SOURCE і SINK — процес, що безпосередньо визначає рівень продуктивності та формування врожаю. Четвертий напрям — пошук фізичного носія «енергетичного стану» клітини. Ним може бути не окрема молекула, а інтегрований структурно-функціональний стан мітохондрій або хлоропластів. І нарешті, розшифрування механізму енергетичного колапсу під час стресу здатне стати ключем до глибшого розуміння природи посухостійкості та розроблення принципово нових підходів до керування адаптивністю рослин.
Таблиця 1
Енергетична модель стресу
| Категорія | Основні положення | Приклади / деталі | Примітки / перспективи |
| Визначення стресу | Стрес як наслідок порушення енергетичного балансу, а не лише як дія несприятливого фактора | Посуха, температура, засолення, дефіцит елементів живлення, патогени | Первинна причина втрати продуктивності — енергетичний дефіцит |
| Ключовий механізм | Дисбаланс між генерацією (фотосинтез, дихання) і споживанням енергії (ріст, репарація, захист) | Енергетичний дефіцит → пригнічення росту, активація захисних механізмів | Системний підхід, об’єднує різні типи стресу |
| Центральна енергетична валюта | АТФ як універсальна «енергетична валюта» | Генерація в хлоропластах та мітохондріях; споживання на підтримку життєдіяльності | Стрес знижує синтез АТФ; активні механізми захисту потребують енергії |
| Енергетичні сенсори | SnRK1 – сенсор дефіциту енергії; TOR – сенсор надлишку енергії | SnRK1 → активує катаболізм і зупинку росту; TOR → стимулює ріст, синтез білка, накопичення біомаси | Баланс TOR / SnRK1 відображає стан енергетичної системи |
| Інтеграція сигналів | Клітина оцінює не лише АТФ, а й комплекс метаболітів, редокс-потік, протонний градієнт, органелярний стан | ATP/ADP, ATP/AMP, NADH/NAD+, ΔΨ, ΔpH, ROS, незгорнуті білки ER та хлоропластів | Первинний сенсор та механізм інтеграції поки не визначені |
| Порогові стани | Рівень енергії визначає ріст, захист або PCD | Високий енергетичний статус → ріст; помірний → захист; критичний → PCD | Гіпотетичний «енергетичний вимикач клітини» |
| Перерозподіл енергії | SOURCE–SINK баланс визначає продуктивність | Перерозподіл ресурсів між ростом та захистом | Механізм поки не детально розкритий |
| Невирішені питання / перспективи | Первинний сенсор дефіциту енергії; інтеграція сигналів; фізичний носій «енергетичного стану»; механізм колапсу | Можливі кандидати: вузли ферментів, органелярні сигнали | Потенціал для революційних відкриттів у біології стресу та селекції |
Нині сучасні моделі енергетичного стресу переважно розглядають абіотичні чинники ізольовано від біотичних впливів, тоді як імунна відповідь описується в межах концепції PTI/ETI без належної інтеграції з метаболічним станом клітини. Натомість запропонована нами інтегрована модель імунітету рослин до патогенів і стресів «SA–JA/ET–ABA–TOR» поєднує сигнальні, енергетичні та метаболічні мережі в єдину регуляторну архітектуру, що визначає рішення рослини щодо росту, захисту або виживання. У центрі цієї мережі функціонує антагоністична вісь TOR ↔ SnRK1 — ключовий енергетичний перемикач, що визначає баланс між анаболізмом і катаболізмом залежно від ресурсного статусу клітини.
TOR активується за умов достатнього вуглецево-азотного забезпечення та позитивного енергетичного балансу, стимулюючи біосинтез білка, клітинний цикл, рибосомогенез і посилення sink strength репродуктивних органів. Натомість за енергетичного дефіциту домінує SnRK1, який ініціює катаболічні програми, індукує автофагію та обмежує енергозатратні процеси росту. Автофагія в цій моделі виступає не лише механізмом рециклінгу макромолекул, а й адаптивним інструментом перерозподілу ресурсів, що забезпечує підтримання C/N балансу та метаболічної гомеостазії під час стресу.
Автофагія постає як універсальна система саморегуляції клітини, що інтегрує контроль енергетичного балансу, якість внутрішньоклітинних структур і підтримання імунної рівноваги. Завдяки керованому розщепленню пошкоджених органел, білкових комплексів і сигнальних молекул вона не лише очищає цитоплазму, а й перенаправляє вивільнені ресурси на ті процеси, які є пріоритетними в умовах стресу. Таким чином, автофагія поєднує функцію «клітинного рециклінгу» з тонким налаштуванням метаболічних потоків.
Водночас її роль виходить далеко за межі метаболічного обслуговування. Автофагія активно модулює імунні реакції, впливаючи на тривалість та інтенсивність гормональних і редокс-сигналів. Через селективну деградацію регуляторних компонентів вона здатна обмежувати надмірну імунну відповідь або, навпаки, підтримувати її на оптимальному рівні. Тому автофагію доцільно розглядати як динамічний регулятор клітинних станів, який формує стабільний баланс між ростом, захистом і відновленням.
У системній стратегії виживання рослин за біотичних та абіотичних стресів автофагія займає ключове місце. Виділяють кілька її форм. Макроавтофагія передбачає утворення аутофагосом, що транспортують великі фрагменти цитоплазми до вакуолі для деградації. Мікроавтофагія здійснюється через безпосереднє інвагінування вакуолярної мембрани з подальшим захопленням невеликих органел або білкових агрегатів. Шаперон-залежна автофагія забезпечує вибіркове спрямування окремих білків до вакуолі за участю молекул-шаперонів, зокрема Hsc70. Кожен із цих механізмів надає клітині гнучкість у відповіді на ушкодження різної природи — від локального окиснювального стресу до системного енергетичного дефіциту. У результаті автофагія координує процеси оборони, регенерації та збереження ростового потенціалу, забезпечуючи адаптивну стабільність організму.
На молекулярному рівні автофагія координується консервативною системою білків ATG, серед яких центральне значення мають ATG5, ATG7 і ATG8. Саме вони забезпечують збирання мембранних структур аутофагосоми та контроль її подальшого злиття з вакуолею. Селективність процесу визначається рецептором NBR1, який розпізнає убіквітиновані білкові агрегати й пошкоджені органели та спрямовує їх до деградації. Ключову роль у запуску мембранної динаміки відіграє PI3-кіназа VPS34, що формує локальні сигнальні домени, необхідні для ініціації аутофагосом. Водночас енергетичні сенсори TOR і SnRK1 виконують інтеграційну функцію: TOR стримує деградаційні процеси за умов достатнього ресурсного забезпечення, тоді як SnRK1 активує автофагію при дефіциті енергії або стресовому навантаженні. Така багаторівнева архітектура дозволяє точно узгоджувати автофагічні процеси з метаболічним і сигнальним станом клітини.
У функціональному вимірі автофагія виступає механізмом очищення й відновлення клітинного середовища. Вона забезпечує видалення ушкоджених мітохондрій, хлоропластів та білкових комплексів, що накопичуються під час стресу, тим самим усуваючи джерела надлишкових активних форм кисню й запобігаючи токсичному перевантаженню метаболітами. У тісній взаємодії з процесами програмованої клітинної смерті (PCD) автофагія може виконувати подвійну роль: з одного боку — стримувати передчасну загибель клітини через зниження редокс-навантаження, з іншого — сприяти локалізованій PCD за умов незворотних ушкоджень. Паралельно вона інтегрується в гормональні мережі, зокрема в JA/ET-, ABA- та SA-залежні сигнальні осі, формуючи адаптивні відповіді на різні типи стресу. Завдяки цьому автофагія підтримує тонкий баланс між захистом, відновленням і збереженням ростового потенціалу — критичну умову довготривалої життєздатності рослини.
Енергетична роль автофагії полягає в її здатності діяти як своєрідний буфер між катаболізмом і анаболізмом. У стресових умовах вона перерозподіляє внутрішні ресурси на користь процесів, необхідних для підтримання базових життєвих функцій, відновлює метаболічну гнучкість і зменшує напруження між витратами енергії на захист та потребами росту. Таким чином, автофагія — це не просто механізм розщеплення й рециклінгу, а інтеграційний центр, що поєднує енергетичні, редокс- і гормональні контури клітини, підтримуючи її стабільність і адаптивну пластичність.
Особливо важливою є її роль у запобіганні надмірній програмованій клітинній смерті (PCD) під час стресу, зокрема при пошкодженні рослин фітофагами. Завдяки вибірковому видаленню ушкоджених органел, білкових агрегатів і джерел надлишкових активних форм кисню автофагія обмежує поширення апоптоподібних сигналів і знижує ризик неконтрольованої втрати клітин. У результаті зберігається функціональна цілісність тканин і підтримується загальна життєздатність рослини, водночас ушкодження локалізуються там, де це найбільш доцільно з точки зору адаптації.
Вибір між автофагією та програмованою клітинною смертю (PCD) залежить від сили й тривалості стресового сигналу. За помірного або короткочасного ушкодження клітина переважно активує автофагію — механізм, що забезпечує видалення пошкоджених структур і відновлення внутрішнього балансу. Якщо ж стрес є інтенсивним або затяжним, можливості автофагії виявляються обмеженими, і запускається PCD. У цьому разі загибель окремих клітин стає адаптивним кроком, який дозволяє локалізувати ушкодження та спрямувати ресурси на підтримку життєздатних тканин. Такий градієнт відповіді — від відновлення до контрольованої елімінації — формує гнучкий механізм адаптивного виживання.
Регуляція PCD тісно інтегрована з редокс-сигналінгом і ATG-білками. Активні форми кисню (ROS) і оксид азоту (NO) виконують роль сигналів, що ініціюють апоптоподібні процеси. Водночас функціонуюча автофагія, опосередкована ATG-комплексами, стримує їх надмірне накопичення, тим самим визначаючи поріг активації PCD. Білки ATG5, ATG7, ATG8 і селективний рецептор NBR1 координують видалення пошкоджених органел і білкових агрегатів — основних джерел оксидативного стресу — та регулюють формування локальних зон клітинної загибелі. У результаті взаємодія автофагії та PCD забезпечує тонкий баланс між локальною руйнацією тканин і збереженням системної стійкості рослини.
У режимах, де домінують сигнальні осі JA/ET, АВА та SA, автофагія й програмована клітинна смерть (PCD) функціонують як взаємопов’язані елементи єдиної адаптивної мережі. Гормональна сигналізація цих шляхів посилює утворення ROS і активацію оборонних ефекторів, водночас індукуючи автофагію як механізм контролю катаболічних і редокс-процесів. Якщо ж ушкодження набуває значної інтенсивності, ті самі сигнали можуть спрямовувати PCD у чітко локалізованих зонах, формуючи стратегічну «жертву» окремих клітин задля збереження функціональної цілісності всього організму.
Енергетична логіка такої стратегії полягає в тому, що часткова елімінація клітин дає змогу перерозподілити ресурси на користь більшої частини тканин і підтримати адаптивну пластичність системи. Автофагія в цьому контексті виступає амортизатором стресу, стримуючи його деструктивний потенціал і запобігаючи масовій загибелі, тоді як PCD реалізується точково й керовано, оптимізуючи співвідношення між обороною та ростом. Така інтегрована архітектура забезпечує динамічну стабільність імунної відповіді та підвищує шанси рослини на виживання за умов ураження фітофагами.
Редокс-сигналізація є додатковим інтегруючим шаром цієї системи. Зміни співвідношення NAD(P)H/NAD(P)+, рівнів реактивних форм кисню (ROS) та антиоксидантного потенціалу виступають швидкими індикаторами клітинного стану. ROS у цьому контексті не лише пошкоджують клітини, а й функціонують як сигнальні молекули, що модулюють активність TOR, SnRK1 та гормональних шляхів SA і JA/ET. Таким чином, редокс-статус безпосередньо впливає на вибір між режимом росту та режимом захисту.
Ключову роль у системному рішенні відіграє мітохондріальний ретроградний сигналінг. Порушення електронтранспортного ланцюга або зміни в дихальній активності ініціюють сигнали до ядра, що перебудовують транскрипційні програми, пов’язані з енергетикою, стресостійкістю та імунітетом. Мітохондрія в цій моделі виступає сенсором енергетичного навантаження та координатором між клітинним метаболізмом і ядерною регуляцією, інтегруючи інформацію про стан дихання, АТФ/АДФ співвідношення та редокс-баланс.
На рівні цілого організму ці процеси проявляються через регуляцію SOURCE–SINK відносин. Перемикання TOR ↔ SnRK1 супроводжується зміною активності транспортерів сахарози (SUT, SWEET), активності інвертаз, фотосинтетичної ефективності SOURCE-органів і сили акцепторних органів (sink strength). Таким чином, сигнально-метаболічний режим визначає напрям і швидкість потоків вуглецю та азоту, перерозподіляючи ресурси між ростом, захистом і виживанням.
Модель представляє рослину як складну багаторівневу систему, у якій сигнальні, енергетичні та метаболічні шляхи функціонують як єдина координаційна мережа, що визначає адаптаційні та захисні реакції. Представлена інтегрована система відкриває можливості для фундаментального розуміння того, як рослини «обчислюють» власний стан і приймають рішення щодо росту, виживання або запуску запрограмованої клітинної смерті.
Цю систему можна описати як динамічний біологічний перемикач, де стан клітини є функцією інтегральних параметрів: F = f(E, C/N, ROS, H), де E — енергетичний статус (АТФ/AMP), C/N — співвідношення вуглецю до азоту, ROS — редокс-потенціал, H — гормональний профіль. Зміна будь-якого з параметрів зсуває систему до альтернативного стабільного стану — ростового (TOR-домінантного) або стресового (SnRK1/ABA-домінантного). Така багатостабільність пояснює, як рослина «обчислює» власний стан і переходить між режимами.
З еволюційної перспективи ці режими сформувалися як оптимальні адаптивні стани під тиском змінного середовища. Вони не є ізольованими реакціями, а представляють інтегровані програми, що дозволяють рослині підтримувати системну рівновагу між продуктивністю та стійкістю. Практично це означає, що кероване модулювання осі TOR ↔ SnRK1, з урахуванням редокс-стану та автофагічної активності, може стати основою нових таргетних технологій управління культурою. Контрольований перехід між ростовим і захисним режимами дозволяє оптимізувати SOURCE–SINK баланс, C/N статус і продуктивність у змінних погодних умовах, відкриваючи шлях до створення справді інтелектуальних антистресових стратегій нового покоління.
Концепція формує наукову основу для створення інноваційних регуляторних технологій вирощування, об’єднаних ідеєю «сигнально-метаболічного управління культурою», та для розроблення таргетних препаратів із чітко визначеними молекулярними мішенями, спрямованих на керовану активацію конкретних фізіологічних режимів. Це можуть бути режими виживання, опосередковані ABA у відповідь на абіотичні стреси, режими імунного захисту на основі SA або JA/ET, а також фаза активного відновлення й росту, залежна від TOR-сигналізації. Ці стани сформувалися у рослин еволюційно як оптимальні адаптивні режими під тривалим тиском абіотичних і біотичних чинників. Вони відображають інтегровані сигнально-метаболічні програми, що забезпечують баланс між виживанням, імунним захистом та продуктивним ростом залежно від умов середовища.
Автофагія постає як ключовий клітинний механізм підтримання гомеостазу, що дедалі виразніше проявляється як інтеграційний вузол між процесами росту, метаболізму та імунної відповіді рослин. У межах JA/ET-, SA- та ABA-опосередкованих типів стійкості вона виконує не допоміжну, а координуючу функцію: забезпечує селективне усунення пошкоджених органел, білкових комплексів і надлишкових сигнальних молекул після дії стресових чинників. Завдяки цьому автофагія розглядається як перспективна біотехнологічна мішень для підвищення толерантності культур до стресів і патогенів без необхідності тривалої або надмірної активації захисних реакцій.
Серед ключових регуляторних ланок протипатогенного і стресовго захисту особливе значення мають гени родини ATG (ATG5, ATG7, ATG8, ATG18), а також сигнальні вузли, що поєднують автофагічні механізми з гормональними та енергетичними контурами клітини. У відповідь на ушкодження, пов’язані з активацією JA/ET-, SA- та ABA-шляхів, індукується селективна автофагія, яка локалізує пошкодження, стримує надмірне накопичення ROS і модулює масштаб програмованої клітинної смерті. У такий спосіб автофагія виступає механізмом «тонкого налаштування» імунної відповіді, забезпечуючи ефективний захист без втрати структурної й функціональної цілісності тканин.
З прикладної перспективи особливу увагу привертають біотехнологічні індуктори автофагії, здатні делікатно активувати цей процес у польових умовах без порушення загального фізіологічного балансу. Одним із провідних агентів такого типу є метилжасмонат, який поєднує запуск JA-залежних захисних програм зі стимуляцією автофагічного потоку, необхідного для перерозподілу та реутилізації ресурсів після стресу. Потенціал мають і ROS-модулятори, що формують контрольований окиснювальний сигнал без розвитку оксидативного колапсу, ініціюючи автофагію як адаптивну відповідь. Окрему категорію становлять інгібітори TOR-сигналізації: послаблюючи ростові програми, вони знімають репресію з автофагічного апарату. У цьому контексті TOR постає не стільки об’єктом пригнічення, скільки регуляторним вузлом, через який можна фазово й дозозалежно переводити клітину зі стану інтенсивного росту в режим адаптації.
Концепція «домінантно-орієнтованих» препаратів ґрунтується на уявленні про рослину як динамічну сигнальну систему, що в кожен момент функціонує в певному регуляторному режимі з домінуванням одного з ключових сигнально-метаболічних контурів — SA, JA/ET, ABA або TOR. Кожна така домінанта визначає не лише характер імунної чи стресової відповіді, а й формує специфічну внутрішньоклітинну біохімічну екосистему, що впливає на розподіл енергетичних ресурсів, гормональні взаємодії, рівень і динаміку ROS, інтенсивність автофагії та загальну метаболічну пластичність клітини.
Розроблення домінантно-орієнтованих препаратів спирається на кілька засадничих принципів. Передусім, кожен продукт має бути спрямований на чітко окреслену молекулярну мішень — NPR1, COI1, SnRK2, TOR або їхні білкові партнери. Водночас необхідно враховувати мережеву природу сигналізації: активація однієї домінанти неминуче модулює інші ланки (зокрема, SA пригнічує JA, ABA знижує активність TOR, а TOR обмежує автофагію). Не менш важливим є узгодження швидкості та тривалості дії: SA і JA/ET забезпечують оперативну відповідь, ABA стабілізує стан системи, тоді як TOR визначає довготривалу програму функціонування. Формуляційна стратегія має будуватися на синергії компонентів, уникаючи їх взаємного антагонізму, а також враховувати агротехнологічні «вікна» застосування — тобто ті фази розвитку або типи стресу, за яких відповідна домінанта є фізіологічно найбільш доречною.
У такій парадигмі створення препаратів, орієнтованих на конкретну сигнальну домінанту, надає агровиробнику інструмент високоточного керування фізіологічним станом рослини, що принципово відрізняється від концепції універсальних біостимуляторів. Цей підхід переводить агросистему з реактивної моделі в керовану, дозволяючи не лише відповідати на вже наявні виклики, а й проактивно визначати сигнальний режим функціонування рослини. У результаті підвищується ефективність усіх складових технології — від захисту та живлення до регуляції росту й реалізації антистресових стратегій. У підсумку автофагія постає як стратегічний компонент інтегрованих систем захисту рослин, який у поєднанні з JA/ET-, SA- та ABA-опосередкованими механізмами формує підґрунтя для біотехнологічно керованої, екологічно виваженої та еволюційно стійкої аграрної практики.
Енергетичний поріг життя: первинний механізм зупинки росту в рослинній клітині.
Найглибше невирішене питання сьогодні у рамках «Енергетичної моделі стресу» — що є первинною причиною зупинки росту? Це фундаментальна діра науки, яка пояснює всю біологію рослини: ріст, стрес, старіння та смерть через єдину систему. Таким чином, енергетична модель стресу існує, але її центральний механізм досі не відкритий і потребує подальших досліджень найвищого рівня.
Сучасна література не дає повної відповіді на ключове фундаментальне питання енергетичного механізму стресу: як клітина «вимірює» свій енергетичний стан і приймає рішення щодо росту, захисту чи програмованої смерті (PCD). Саме ця прогалина відкриває потенціал для революційних відкриттів у біології стресу.
Наша концепція, «Universal Energy Threshold Model of Plant Stress» і «Energy Allocation Model Explains Yield Loss», у контексті «Інтегративнї моделі імунітету рослин від патогенів і стресів» пояснює, як клітина «вимірює» свій енергетичний стан і приймає рішення про ріст або виживання. Вона передбачає існування критичного енергетичного порогу, нижче якого ріст клітини припиняється, а активуються механізми виживання. Зниження врожайності при цьому виникає не через пряме пошкодження тканин, а через стратегічний перерозподіл енергії. Ця концепція пропонує революційний підхід, акцентуючи увагу на первинному енергетичному сигналі, а не на гормонах чи генах.

Фото 3. Стрес як енергетична криза.
Згідно моделі, первинними маркерами енергетичного стану клітини є комбінація класичних метаболітів, редокс-потоку та органелярних сигналів. Серед кандидатів найпоширеніші: співвідношення ATP/ADP та AMP, яке активує сенсорні кінази SnRK1/AMPK; редокс-пара NAD(P)H/NAD(P)+, що відображає баланс окислення/відновлення і метаболічну активність; протонний градієнт на мембранах мітохондрій та хлоропластів (ΔΨ, ΔpH), який інтегрує роботу електронного транспорту і потенціал енергетичної ємності; а також органелярні сенсори стресу, наприклад накопичення незгорнутих білків у хлоропласті чи ендоплазматичному ретикулумі, що сигналізує про перевантаження енергетичних ресурсів і може запускати захисні програми або PCD. Таким чином, первинний маркер — це не один метаболіт, а інтегрована комбінація енергетичних та органелярних сигналів.
Перехід між домінантними станами відбувається не випадково, а відповідно до принципу «вікон чутливості», які задають оптимальні часові та функціональні інтервали для активації чи пригнічення певної програми. Ці вікна формуються під впливом енергетичного статусу рослини, сили та тривалості стресового сигналу, фази онтогенезу й наявної імунної пам’яті. Так, SA-домінанта може повноцінно реалізуватися лише за умови тимчасового гальмування TOR-залежних ростових програм і готовності JA/ET-сигналізації до взаємодії. Подібним чином, після завершення гострої фази стресу ABA-опосередкована стабілізація поступово змінюється TOR-домінантою, що ініціює відновлення росту та акумуляцію біомаси.
Жодна домінанта не функціонує ізольовано — вона реалізується в межах специфічного «вікна чутливості», тобто комплексу умов, за яких може стати провідною в мультистабільній сигнальній мережі. Саме це пояснює, чому одна й та сама сигнальна молекула або стресовий чинник здатні викликати різні реакції залежно від контексту, енергетичного балансу клітини та стадії розвитку.
Визначальним інтегратором такого вибору виступає рівень активних форм кисню (ROS). Низькі концентрації ROS підтримують TOR-орієнтовану стабілізацію та перевагу анаболічних, ростових процесів. Помірні рівні ROS створюють умови для активації JA/ET-залежних захисних відповідей, що є критично важливим при атаках некротрофних патогенів або фітофагів. Високі ж концентрації ROS зсувають баланс у бік SA-домінанти, стимулюючи антибіотроф-спрямовану імунну реакцію. Отже, ROS-вікно виконує роль сигнального інтегратора, який визначає домінантний стан системи залежно від інтенсивності оксидативного навантаження.
Поріг енергії (енергетичне вікно (ATP/AMP)), що визначає перемикання між ростом, захистом і PCD, можна описати наступним чином. Високий енергетичний статус (високий ATP/NADPH, стабільний ΔΨ) дозволяє клітині зростати і ділитися. Помірне зниження енергетичних ресурсів активує захисні програми, включно з антиоксидантною відповіддю, аутофагією та збереженням ресурсів. Різке падіння енергії або критичний редокс-стрес запускають програмовану смерть клітини, щоб запобігти поширенню пошкоджених клітин у тканині. Порогова логіка є мультикомпонентною й інтегративною: вона залежить від типу клітини та її поточного стану, тому не зводиться до простого значення ATP.
Енергетичний стан клітини відіграє ключову роль у визначенні провідної домінанти. Високий рівень ATP стимулює активність TOR, що підтримує ростові процеси, одночасно пригнічуючи ABA та дозволяючи JA/ET-відповідям ефективно функціонувати під час фази відновлення. У протилежному випадку, при низькому енергетичному статусі або підвищенні AMP, активуються SnRK1 і SnRK2, що запускають ABA-орієнтовані каскади та гальмують JA/ET-сигналізацію, переводячи систему у режим виживання. Таким чином, співвідношення ATP/AMP визначає стратегічний вибір клітини — спрямування ресурсів на ріст або активацію захисних програм.
Гормональні взаємодії створюють додаткові пороги чутливості між домінантами, утворюючи так звані «гормональні вікна» (SA/JA/ABA). Підвищення SA піднімає поріг для JA, запобігаючи одночасній конфліктній активації цих шляхів. Навпаки, JA/ET знижують поріг SA-активації, забезпечуючи швидку реалізацію JA-домінанти у відповідь на некротрофні патогени або фітофагів. ABA підвищує чутливість до стресових сигналів навіть за низьких рівнів SA та JA, дозволяючи системі оперативно реагувати на абіотичні фактори та зміщувати баланс у бік виживання під час енергетичного дефіциту.
Фазовість сигналів визначає як кінцеву відповідь, так і часові «вікна» для активації домінант. Одні й ті самі ROS чи гормональні сигнали можуть спричиняти різні реакції залежно від стадії інфекції або тривалості стресу. Наприклад, ранні піки ROS здебільшого активують JA/ET-домінанту, тоді як пізні — SA-домінанту. У випадку тривалого стресу або значного зниження енергетичного стану перевага переходить до ABA, що забезпечує адаптацію та виживання клітин.
Отже, домінанти не просто конкурують між собою; їхня реалізація залежить від того, наскільки молекулярні перемикачі активуються у відповідних «вікнах чутливості». Влучне «попадання» у ROS-, енергетичне, гормональне та часові вікна визначає, яка домінанта переважатиме, формуючи фазову, контекстно-залежну адаптацію рослини до біотичних та абіотичних викликів.
Мережа імунітету та метаболізму рослин має мультистабільну природу, що передбачає, що домінанти не діють ізольовано, а постійно взаємодіють і змагаються за пріоритет у використанні обмежених ресурсів. Ця конкуренція реалізується через ключові молекулярні перемикачі, які координують одночасну активацію та пригнічення різних домінантних станів. До основних таких регуляторів належать SnRK2-кінази, TOR/mTOR-кінази, NPR1 та комплекс COI1-JAZ. SnRK2 керує ABA-залежною адаптацією до водного та абіотичного стресу, пригнічуючи паралельні ростові або імунні програми. TOR, навпаки, інтегрує сигнали енергетичної достатності та поживного забезпечення, стимулюючи ріст і біосинтез, водночас обмежуючи активацію SA- та JA/ET-домінант. NPR1 виступає ключовим координатором SA-домінанти, забезпечуючи системну резистентність та пригнічення біотрофних шляхів, тоді як комплекс COI1-JAZ визначає швидкість і силу JA/ET-відповіді на ушкодження тканин і атаку комах та некротрофних патогенів.
Інтеграція всіх енергетичних сигналів у єдине рішення клітини здійснюється через центральні вузли, такі як SnRK1, TOR та GCN2, які об’єднують метаболіти, редокс-потік та гормональні сигнали. Органели — мітохондрії, хлоропласти та ER — генерують локальні сигнали (ROS, незгорнуті білки), які передаються до ядра. Клітина реалізує порогову мультистабільність: вона оцінює комбінацію енергетичних, редокс- та стресових сигналів і приймає одне з трьох рішень — ріст, захист або PCD. Це можна уявити як інтегратор, який обчислює «енергетичний вектор» клітини і запускає відповідну програму.
Молекулярні перемикачі виступають ключовими регуляторами домінантних станів рослин, підтримуючи тонкий баланс між ростом, імунною активністю та адаптацією до стресових умов. Саме вони визначають, яка домінанта — ABA, SA, JA/ET або TOR — стає провідною за конкретних зовнішніх і внутрішніх обставин.
SnRK2 є центральним перемикачем ABA-домінанти та сигналів енергетичного дефіциту. Його активація відбувається під час абіотичних стресів — таких як нестача води, холод чи гербіцидне пошкодження — а також при накопиченні ABA та зниженні рівня енергетичних метаболітів (ATP/AMP). Після запуску SnRK2 активуються ABA-орієнтовані сигнальні каскади через транскриптори ABF/AREB, пригнічується ріст, інгібується JA/ET-сигналізація і послаблюється TOR-активність. Це зміщує клітинну систему у режим виживання, дозволяючи ефективно протистояти абіотичним викликам. Таким чином, SnRK2 слугує ключовим перемикачем, який переводить домінанту з JA/ET у ABA-орієнтований стан.
TOR (Target of Rapamycin) виступає основним перемикачем росту та антистресової стабілізації. Він активується при високому енергетичному статусі клітини (високий ATP/AMP) та достатньому забезпеченні поживними речовинами. TOR підтримує біосинтетичні програми росту, пригнічує автофагію, стабілізує JA/ET-відповіді у фазі відновлення та частково інгібує SnRK1 і SnRK2. Активність TOR утруднює перехід клітини в SA- або ABA-домінанту, тим самим виступаючи ключовим перемикачем у моделі «ріст/захист» і забезпечуючи ефективне відновлення продуктивності рослини після стресу.
NPR1 є центральним регулятором SA-домінанти. Його активація відбувається у відповідь на підвищені рівні ROS та саліцилової кислоти (SA). NPR1 стимулює експресію SA-залежних генів, таких як PR1 та PR2, пригнічує JA-шлях через взаємодію з TGA- та WRKY-транскрипторами, зміщуючи систему від JA/ET до SA, а також частково інгібує TOR через активацію стресових програм. Таким чином, NPR1 формує антибіотроф-орієнтований імунний режим і координує системну набути стійкість.
COI1-JAZ виступає ключовим перемикачем JA/ET-домінанти. COI1 функціонує як рецептор гормону JA-Ile: при його зв’язуванні відбувається деградація репресорів JAZ, активація транскрипторів MYC2, MYC3 та MYC4, пригнічення SA-сигналізації та стабілізація ET-блоку (EIN2/EIN3). Завдяки цьому COI1-JAZ забезпечує швидку і точну активацію JA/ET-домінанти, що особливо важливо під час атак некротрофних патогенів або механічних ушкоджень, коли ріст та SA-відповіді тимчасово повинні бути обмежені.
Таким чином, ці молекулярні перемикачі працюють як координаційна система, яка динамічно змінює домінанти у рослині, формуючи багатофазну адаптацію до стресів і патогенів. SnRK2 керує ABA-орієнтованими стратегіями виживання, TOR відповідає за відновлення росту, NPR1 визначає SA-орієнтовану імунну відповідь, а COI1-JAZ спрямовує JA/ET-захисні програми, створюючи інтегровану мережу адаптаційних реакцій.
Отже, сучасна концепція енергетичного стресу у контексті «Інтегративної моделі імунітету рослин від патогенів і стресів» в рослинних клітинах виглядає так: первинні маркери — ATP/AMP, NAD(P)H та органелярний стан; пороги — високий енергетичний статус → ріст, помірний → захист, критичний → PCD; інтеграція сигналів здійснюється через SnRK1/TOR та органелярні сигнали, що формують єдину мультистабільну відповідь. Ця модель дає пряму відповідь на фундаментальне питання енергетичного механізму стресу — як клітина «вимірює» свій енергетичний стан і приймає рішення щодо росту, захисту чи зупинки. Вона підкреслює роль енергетичних сенсорів (SnRK1, TOR, GCN2) та перерозподіл ресурсів (метаболіти, цукри, амінокислоти), але це лише частина складної системи сигналів. Модель також окреслює, який сигнал може виступати первинним «енергетичним маркером» клітини, як інтегруються всі енергетичні сигнали в єдине рішення та який критичний поріг енергії запускає захисні стратегії або PCD.
Сучасна теорія клітинного стресу вважає, що основним сигналом енергетичного дефіциту є падіння ATP. У рамках інтегративної моделі імунітету рослин до патогенів і стресів «SA–JA/ET–ABA–TOR» первинними маркерами енергетичного стану клітини є комбінація класичних метаболітів, редокс-потоку та органелярних сигналів (ATP/AMP, NAD(P)H та органелярний стан), а головним енергетичним сигналом може також бути ферментна нестабільність як первинний енергетичний сенсор. Зупинка росту спричиняється не самим виснаженням АТФ, а нестабільністю ключових енергочутливих ферментативних вузлів, що призводить до колапсу метаболічного потоку нижче критичного системного порогу. Клітина не вимірює рівень АТФ напряму. Вона “відчуває”, що мережа почала руйнуватися. І руйнування починається в конкретних вузлових ферментах. Первинним енергетичним сигналом є втрата структурно-кінетичної стабільності невеликої групи енергозалежних “вузлових” ферментів, і саме їхня деградація активності запускає перемикання між ростом і виживанням.
Якщо критичний тригер — це не ATP, а втрата стабільності вузлових ферментів, тоді наслідки будуть такими: теорія стресу зміниться, адже стрес слід розглядати як мережевий колапс; теорія старіння зміниться, бо старіння стає наслідком накопичення нестабільності ферментних вузлів; теорія росту зміниться, оскільки ріст визначається стабільністю ферментного потоку; і, нарешті, селекція зміниться, орієнтуючись на відбір за стабільністю ключових ферментів, а не лише за рівнем їхньої експресії.
Сьогодні є серйозні експериментальні дані про значення стабільності окремих ферментів (наприклад, Rubisco Activase) у визначенні того, як організм реагує на стрес, часто незалежно від глобального рівня ATP. Однак загальна концепція того, що саме ферментна нестабільність замість ATP є первинним універсальним сенсором клітини, ще не закріплена в літературі. Вона має підстави для дослідження, і частина окремих результатів її підтримує, але наразі це — перспективна гіпотеза.
Таблиця 2
Логіка порогів переключення сигнально-енергетично-метаболічної захисної програми у відповідний стабільний стан у контексті інтегрованої моделі імунітету рослин від патогенів і стресів «SA–JA/ET–ABA–TOR»
| Рівень | Компоненти | Функція | Порогова відповідь | Примітки |
| Енергетичний маркер | ATP/ADP/AMP, NAD(P)H/NAD(P)+, ΔΨ/ΔpH | Відображають загальний енергетичний стан та метаболічну активність клітини | Високий енергетичний статус → ріст; помірний дефіцит → захист; критичний дефіцит → PCD | Первинний маркер – інтеграція рівня АТФ, редокс-потоку та органелярного стану |
| Органелярні сигнали | Мітохондрії, хлоропласти, ER; ROS, накопичення unfolded proteins | Локальні сигнали перевантаження та стресу, передаються до ядра | Допомагають визначити порогове рішення: ріст/захист/PCD | Інтегрують локальні стресові умови з глобальним енергетичним станом |
| Центральні сенсори | SnRK1, TOR, GCN2 | SnRK1 активується при енергетичному дефіциті, стимулює катаболізм і виживання; TOR реагує на достатній енергетичний ресурс, стимулює ріст і синтез білка; GCN2 контролює амінокислотний та стресовий стан | Мультистабільне рішення: ріст / захист / PCD | Інтегрують метаболіти, редокс-потік та гормональні сигнали |
| Гормональні шляхи захисту від біотичних і абіотичних стресів | SA (саліцилова кислота), JA/ET (жасмонова кислота/етилен), ABA (абсцизова кислота) | Координують відповідь на біотичні та абіотичні стреси, взаємодіють із енергетичними сенсорами | Активують захисні механізми при помірному дефіциті енергії | Взаємодія з SnRK1/TOR посилює адаптивну відповідь |
| Гормональні шляхи росту | TOR, цитокініни, гібереліни, ауксини | Стимулюють ріст, анаболізм, фотосинтез, накопичення ресурсів | Активується при достатньому енергетичному та вуглецево-азотному статусі; пригнічує SnRK1 | Підвищує активність транспортерів SUT/SWEET та ферментів cell wall invertase, посилюючи sink strength |
| Мережеві ефекти / вузлові ферменти | Ключові енергозалежні ферменти (наприклад, Rubisco Activase) | Ферментна нестабільність виступає первинним сигналом енергетичного дефіциту | Втрата стабільності вузлових ферментів → колапс метаболічного потоку → перемикання між ростом і виживанням | Підкреслює гіпотезу ферментної нестабільності як первинного тригера |
| Інтеграція сигналів / рішення клітини | SnRK1/TOR + органелярні сигнали + гормони | Обчислюють «енергетичний вектор» клітини та запускають відповідну програму | Високий енергетичний статус → ріст; середній → захист; критичний → PCD | Порогова мультистабільність залежить від типу клітини та її стану |
| Функціональний результат | Ріст / Захист / PCD | Визначає подальший розвиток рослини під стресом | Активується стратегічний перерозподіл енергії між SOURCE і SINK | Зниження врожайності виникає через енергетичний перерозподіл, а не пряме ушкодження тканин |
У практичному застосуванні ця інтегрована модель відкриває можливість стратегічно синхронізувати домінанти в агрономічних рішеннях. Планування використання антистресантів, біостимуляторів, гормональних індукторів та мікробіомних препаратів може бути спрямоване на оптимальне поєднання фаз чутливості рослини з активацією потрібного домінантного стану. Так, ABA-індуктори доцільно вводити під час гострих абіотичних стресів, тоді як TOR-активатори ефективні після стабілізації стану рослини, забезпечуючи швидке відновлення росту та продуктивності. Координація SA- та JA/ET-домінант у періоди високого патогенного ризику дозволяє уникнути конфліктів між сигналами та підвищує ефективність захисних реакцій проти різних патогенів і фітофагів.
Глибоке розуміння механізмів переключення гормональних домінант відкриває можливості для точного керування рослиною за допомогою технологічних і біостимуляторних підходів. Агроінтервенції можна проєктувати так, щоб активувати потрібну домінанту у відповідному «вікні чутливості», зменшуючи ризик небажаних переходів і підвищуючи адаптивність до конкретних стресових факторів.
Контроль над рівнем ROS дозволяє регулювати баланс між ростом та імунною відповіддю, керуючи ROS-вікном. Використання антиоксидантів допомагає знизити надмірну активність SA-домінанти та запобігає небажаному домінуванню антибіотроф-орієнтованого стану при незначному стресі. Навпаки, легкі ROS-індуктори можуть стимулювати JA/ET-домінанту, що ефективно протидіє некротрофним патогенам і фітофагам, особливо на ранніх етапах інфекції.
Підтримка високого енергетичного рівня є критичною для активації TOR і запуску ростових програм після стресових впливів, забезпечуючи контроль над енергетичним балансом клітини (ATP/AMP). Біостимулятори, збагачені амінокислотами, джерелами азоту або метаболітами, що покращують мітохондріальне дихання, сприяють стабілізації TOR і JA/ET-домінанти у фазі відновлення, запобігаючи небажаному переходу системи до ABA-домінанти.
Активне регулювання автофагії за допомогою лактату та азотовмісних метаболітів підтримує очищення клітин від надлишку ROS, перешкоджаючи неконтрольованому зміщенню з JA/ET у ABA-домінанту. Це дозволяє забезпечити збалансовану імунну відповідь без втрати ростового потенціалу.
Застосування «часових вікон» у введенні препаратів дозволяє ефективно синхронізувати домінанти з фазою стресу або інфекції. Ранні JA/ET-індуктори орієнтовані на боротьбу з некротрофами, тоді як пізні SA-індуктори активують антибіотроф-орієнтовані механізми. ABA-модулятори доцільно застосовувати при тривалих абіотичних стресах, таких як посуха або холод, для підтримки режиму виживання та активації SnRK2-сигналізації.
Практична реалізація синхронізації домінант передбачає поєднання антистресантів, біостимуляторів та технологічних заходів: після гербіцидного стресу рекомендується стимулювати TOR-комплекс разом з антиоксидантами, щоб уникнути небажаного переходу до SA-домінанти; у разі загрози некротрофних патогенів застосовується JA/ET-комплекс у «середньому ROS-вікні»; при холоді або дефіциті води робиться акцент на підтримці ABA-домінанти та активності SnRK2.
Таким чином, інтегрована модель демонструє, що домінанти конкурують і перемикаються через молекулярні перемикачі у визначених вікнах чутливості, а грамотне використання цього принципу в агрономічній практиці дозволяє прогнозовано керувати імунітетом, стресостійкістю та продуктивністю рослин.
Таблиця 3
Як домінанти змагаються і переключаються
| Молекулярний перемикач | Відповідна домінанта | Біохімічний ефект | Фаза / «вікно чутливості» | Приклад застосування |
| SnRK2 | ABA | Активація адаптації до абіотичного стресу, закриття продихів, енергозбереження | Гострий абіотичний стрес (посуха, холод, гербіциди) | ABA-індуктори під час стресу |
| TOR / mTOR | TOR | Активація росту та біосинтезу, пригнічення SA/JA | Після стабілізації після стресу | TOR-активатори для відновлення продуктивності |
| NPR1 | SA | Індукція системної набутої резистентності, PR-білки, контроль біотрофів | Високий патогенний ризик (біотрофи, віруси, бактерії) | SA-індуктори або біостимулятори проти біотрофів |
| COI1-JAZ | JA/ET | Захист від некротрофів та фітофагів, синтез протеазних інгібіторів | Ураження тканин або атака комах | JA-індуктори при механічних ушкодженнях або шкідниках |
| Комбінації SnRK2+TOR, SA+TOR, JA+TOR | ABA/TOR, SA/TOR, JA/TOR | Модулюють баланс росту і захисту | Переходи між стресовою стабілізацією та ростом | Фазове застосування біостимуляторів, гормонів або мікробіомних препаратів |
Молекулярно-клітинні і фізіологічні основи стресу рослин.
На початкових етапах розвитку стресу його неможливо визначити візуально, оскільки рослина зовні ще не відрізняється від здорової. У цей період ключові зміни відбуваються приховано — на клітинному та молекулярному рівнях. Вони пов’язані з глибокою перебудовою метаболізму, коли рослина перемикається з програм росту на програми захисту. Ці два функціональні режими є фізіологічно антагоністичними: процеси росту спрямовані на синтез біомаси та формування врожаю, тоді як захисні реакції спрямовані на виживання і потребують значних витрат енергії, що тимчасово обмежує продуктивність.
Візуальні симптоми з’являються значно пізніше, коли стрес уже досягає глибокої стадії та супроводжується суттєвими порушеннями клітинних структур. Це проявляється зміною забарвлення листків, пригніченням фотосинтезу, пошкодженням хлоропластів і розвитком частково або повністю незворотних процесів у тканинах. На цьому етапі рослина вже зазнала фізіологічних втрат, які безпосередньо знижують реалізацію її врожайного потенціалу.
Особливо важливо, що кожен окремий стресовий епізод протягом вегетації залишає свій кумулятивний ефект. Такі стреси накопичуються, формуючи так званий «сумарний стрес сезону», який може зменшувати врожайність культури на 60 % і більше. Це пов’язано з тим, що навіть короткочасне пригнічення фотосинтезу, росту або енергетичного обміну має довготривалі наслідки для формування генеративних органів і кінцевої продуктивності.
У зв’язку з цим особливого значення набуває розробка та впровадження інструментальних методів ранньої діагностики стресу рослин. Саме раннє виявлення фізіологічних порушень — ще до появи видимих симптомів — відкриває можливість своєчасного втручання, стабілізації клітинного метаболізму, збереження функції фотосинтетичного апарату і, як наслідок, мінімізації втрат урожайності. Такий підхід закладає основу сучасної превентивної агрономії та регуляторних технологій вирощування культур, у яких центральне місце займає управління біологічними режимами рослини, а не механічна компенсація окремих стресових факторів. У цій системі контроль продуктивності починається ще на стадії прихованих клітинно-метаболічних змін, задовго до появи видимих симптомів пошкодження.
На клітинному рівні будь-який стрес супроводжується розвитком комплексу взаємопов’язаних процесів, серед яких ключовими є:
– пригнічення фотосинтезу;
– накопичення реактивних форм кисню (ROS);
– пошкодження мембран;
– деградація білків;
– порушення іонного балансу.
Особливо важливим є пошкодження хлоропластів, які є основним джерелом енергії в клітині. Саме фотосинтез забезпечує синтез АТФ та відновлювальних еквівалентів, необхідних для росту і формування врожаю. При стресі ефективність фотосинтезу може знижуватися на 30–80%, що призводить до різкого дефіциту енергії.
У відповідь рослина активує захисні механізми, включаючи:
– синтез антиоксидантів;
– продукцію стресових білків;
– автофагію пошкоджених структур;
– синтез осмопротекторів.
Абіотичні стреси активують складні гормональні, редокс-сигнальні та транскрипційні мережі у рослинах. Різні типи стресів мають частково спільні молекулярні елементи (ROS, ABA, TFs), але також специфічні відповіді (HSPs при теплі, дегідрини при холоді тощо), що вимагає діференційованого підходу при застосуванні антистресових речовин. Так, за умов посухи стратегія має бути спрямована на підтримання водного гомеостазу та регуляцію транспірації — оптимальним є використання осмопротектантів у поєднанні з ABA-агоністами, що дозволяє стабілізувати тургор клітин і зберегти SOURCE–SINK баланс. При засоленні ґрунтів ключовим є контроль іонного гомеостазу: акцент переноситься на підтримання співвідношення Na⁺/K⁺ через регуляцію HKT- та SOS-систем, доповнену потужним антиоксидантним захистом для зменшення вторинного оксидативного стресу. За дії заморозків першочерговим стає збереження структурної цілісності клітини — мембранна стабілізація та індукція LEA-білків забезпечують зниження кристалізаційного пошкодження й підтримку ферментативної активності. В умовах високих температур стратегія має бути спрямована на протеомну стабільність і фотосинтетичний захист: індукція HSP у поєднанні із захистом фотосистеми II дозволяє зменшити денатурацію білків та фотоокислювальний стрес. За підвищеної кислотності ґрунтів основною мішенню є детоксикація Al³⁺ та підтримання функціональної стабільності кореневої системи, що забезпечує збереження поглинальної здатності та метаболічної активності. Нарешті, при гербіцидному стресі пріоритетною є активація систем детоксикації, зокрема глутатіонової ланки, що дозволяє мінімізувати ушкодження метаболічних шляхів і зберегти енергетичний потенціал клітини.
Таким чином, ефективна антистресова стратегія повинна бути таргетованою — орієнтованою на конкретний механізм пошкодження та відповідні молекулярні вузли регуляції. Проте всі ці процеси потребують значних енергетичних витрат. Тому формується фундаментальний конфлікт між ростом і виживанням. Для зняття стресу у рослин антистресові препарати повинні містити комбіновані антистресові формуляції (осмопротектант + антиоксидант + гормональний модуль, що підхдить за типом стресу). Важлива також регуляція SOURCE–SINK балансу через підтримку фотосинтезу та транспорту асимілянтів.
Таблиця 4
Узагальнення: стратегія стабілізації енергетичного гомеостазу від абіотичного стресу
| Стрес | Ключовий акцент |
| Посуха | Осморегуляція + ABA |
| Засолення | Na⁺/K⁺ баланс + антиоксиданти |
| Заморозки | Мембранна стабілізація + LEA |
| Спека | HSP + захист PSII |
| Кислотність | Детоксикація Al³⁺ + коренева стабільність |
| Гербіциди | Детоксикація + глутатіонова система |
Таблиця 5
Систематизація антистресових сполук за типами абіотичного стресу
| Тип стресу | Механізм пошкодження | Маркери стресу | Ключові молекулярні вузли | Ефективні антистресові сполуки | Агрономічні рекомендації |
| Посуха | Осмотичне зневоднення, ↓тургор, порушення фотосинтезу, накопичення ROS | ↑ABA, ↑ROS, ↑пролін/β-трейн, ↓Fv/Fm | ABA–PYR/PYL–SnRK2, DREB/AREB, антиоксидантні ферменти | Пролін, β-трейн, мелатонін, амінокислоти, гумати | Позакореневе внесення у фазу кущення/бутонізації; превентивна обробка за 2–3 дні до прогнозованої посухи. |
| Тепловий стрес | Денатурація білків, порушення мембран, пригнічення фотосинтезу, ROS | ↑HSPs, ↑ROS, ↑MDA, тепловий хлороз | HSF–HSP система, антиоксидантний каскад, Ca²⁺-сигналінг | Мелатонін, брассиністероїди, аскорбат, амінокислоти | Обробка за 24 год до спеки; ранкові обприскування; поєднання антиоксидантів з біостимуляторами. |
| Холод/заморозки | Ригідність мембран, розриви мембран льодом, ROS | ↑CBF/DREB, ↑LEA/Dehydrin, ↑пероксидазна активність | ICE1–CBF–COR каскад, ABA-сигналінг, антиоксидантна система | Мелатонін, амінокислоти, білкові гідролізати, фульвати | Превентивне внесення за 24–72 год до заморозків; повторна обробка після стресу для відновлення. |
| Засолення | Осмотичний стрес, Na⁺/Cl⁻ токсичність, іонний дисбаланс, ROS | ↑Na⁺ у тканинах, ↑пролін, ↑ABA, ↓фотосинтез | SOS-путь (SOS1/2/3), HKT-транспортери, антиоксидантні ферменти | Гліцин-бетаїн, пролін, мелатонін, кремній, гумати | Кореневе внесення (пролонговане) на ранніх фазах; позакореневі обробки при перших ознаках іонного стресу. |
| Кислотність ґрунту (Al³⁺) | Токсичність Al³⁺ → пошкодження кореня, ↓всмоктування P/Ca/Mg | ↑органічні кислоти (малат, цитрат), пригнічення росту кореня | ALMT-транспортери, регуляція pH ризосфери | Кальцій, кремній, гумати, мікробні інокулянти | Вапнування ґрунту; внесення Ca/Si у зону кореня; локальне внесення біопрепаратів (Al-tolerant бактерії). |
| Гербіцидний стрес | Інгібіція фотосистем, блокада ферментів ALS/EPSPS, окиснювальний стрес | ↑ROS, ↑MDA, пригнічення росту | Антиоксидантна система, детокс. ферменти (GST, P450) | Амінокислоти, глутатіон, гумати, екстракти водоростей | Обробка через 3–5 днів після гербіциду; поєднання з антиоксидантами. |
Біотичний стрес викликається живими агентами – грибковими патогенами, бактеріями, вірусами, фітонематодами, шкідниками – і призводить до значних втрат врожаю (20–40 % світового виробництва). Рослини мають складну систему захисту (PTI/ETI, гіперчутливу відповідь, системний накопичувальний/індукований імунітет), що регулюється сигнальними гормонами: саліциловою кислотою (SA) – проти біотрофних патогенів, та осі JA/ЕТ – проти некротрофів та фітофагів. З метою посилення стійкості застосовують біоактивні речовини (мелатонін, SA, JA, пептиди) і біопрепарати (Trichoderma, Bacillus тощо), а також агротехнічні прийоми (сівозміна, обприскування на конкретних фазах росту та ін.).
Таблиця 6
Антистресові сполуки за типами біотичного стресу
| Тип стресу | Механізм пошкодження | Маркери стресу | Ключові вузли (молекули) | Антистресові речовини | Агрономічні стратегії (обробки) |
| Гриби | Розповсюдження гіфів, некрози, гнилі; плямистість, в’янення | ROS (H₂O₂, O₂⁻), MDA; PR-білки (β-1,3-глюканаза, хітиноза), фітоалексини | SA (проти біотрофів) та JA/ЕТ (проти некротрофів); WRKY/TGA; PR-білки (PR1, PR2); ROS, фітоалексини | Мелатонін (антиоксидант, індуктор PR); SA/BTH (SAR-індуктори); JA/метил-JA; хітозан; Trichoderma/Bacillus (біоконтроль) | Протруювання насіння Trichoderma/Bacillus (стимуляція імунітету); обприскування на стадії сходів або перед квітненням; SA/мелатонін як превентивні обробки |
| Бактерії | Проникнення через рани/продихи; заселення міжклітин; токсини, гідролітичні ферменти; в’янення, пухлини | ROS, підвищена пероксидазна активність; PR-білки (PR1–PR5); фенольні сполуки; запізній втягнення води | SA (SAR) як головний сигнал; WRKY, MYB, NAC TFs; NPR1; NO; PR-білки | Мелатонін (бактерицидний, індукує резистентність); SA (індуктор SAR); Bacillus/Biocom (антибіотики і ISR); ABA у деяких випадках | Полив/протруювання Bacillus; листкові обприскування SA (приблизно вегетація 2–4 листки); контроль резервуарів інфекції (видалення хворих тканин) |
| Віруси | Внутрішньоклітинна реплікація; системне поширення; викликають мозаїку, хлороз, карликовість | ROS (H₂O₂, .NO); PR-білки (особливо PR1, PR10, рибонуклеази); гіперіндукція генів SAR; специфічна антивірусна РНК | SA – центральний гормон (SAR); WRKY- та TGA-фактори; NB-LRR- білки (R-білки); PR10, RNS (рибонуклеази) | Мелатонін (скорочує вірусну РНК, знижує концентрацію вірусу); SA (системна резистентність); JA/ЕТ (у деяких випадках); RNAi-дсРНК; ендофіти Trichoderma | Використання чистого насіння (вірус-фрі); контроль переносників (проти сокосисних комах); превентивні обприскування SA/BTH перед сезоном; підтримка високого стану рослин (мінерали, волога) |
| Нематоди | Інвазія коренів, формування вузлів/галлів; висмоктування клітинного соку (підвищене утворення галлів → кущіння, в’янення) | Галлі в коренях; підвищення JA/ЕТ; ROS (H₂O₂); активність фенілпропаноїдних ферментів; PR-білки (особливо β-1,3-глюканази) | JA/ЕТ (LOX, AOS); TFs WRKY, MYB, NAC; PAL (фенілаланінамоніаз); в системі захисту NAC і Jasmonate-зв’язані TF | Bacillus spp. (в т. ч. B. velezensis – нематоцидний, індукує WRKY, PAL, PR); Trichoderma spp.; бурштинова кислота; рослинні екстракти (репеленти) | Сівозміна (несприятливі культури), ґрунтові внесення Bacillus/Trichoderma перед посівом; вирощування уловлювачів nematodes; обробка комбінаціями (Bacillus + стимулятор росту) у фазі сходів |
| Шкідники | Гризучі – знищують тканини листя/плодів; сисні – висмоктують соки, переносять віруси | JA/ЕТ-відповіді; водорозчинні феноли; індукція протеазних інгібіторів; ROS (високий H₂O₂); скидання легких летких органічних сполук (VOC); MDA (перекисне ушкодження мембран) | JA (MYC2, COI1-рецептор); системін/інцептин-пептиди; TFs MYC, bHLH, WRKY; SA (у відповідь на кліщів); PR-білки PI-II (інгібітори протеаз) | Мелатонін (підвищує феноли/алкалоїди, модулює JA); JA/метил-JA; Bacillus thuringiensis (Bt) проти гусениць; Beauveria spp., Metarhizium spp.; піретроїди/спіносад; рослинні репеленти (нікол, терпени) | Інтегрована захист (ентомофаги, феромонні пастки); обприскування JA/SA при появі перших ознак ушкодження; профілактична обробка біоензимами (збуджуючими захист); кондиціонування рослин перед сезоном шкідників |
Будь-яка стресова програма для рослини є значно енергозатратною, оскільки активація захисних механізмів потребує мобілізації додаткових метаболічних ресурсів, що неминуче формує компроміс між ростом і захистом. При нормальних умовах при рості рослини більшість енергії використовується для анаболічних процесів — синтезу білків, клітинних структур і формування біомаси. Проте за дії стресу — посухи, холоду, високих температур, патогенів або хімічних факторів — значна частина ресурсів перенаправляється на підтримання клітинної цілісності, детоксикацію, ремонт пошкоджень і активацію захисних систем. Центральну роль у координації цього балансу відіграють ключові сенсори енергетичного стану клітини, серед яких SnRK1 (SNF1-related kinase 1), TOR (Target of Rapamycin) і GCN2 (General Control Nonderepressible 2). SnRK1 функціонує як універсальний сенсор енергетичного дефіциту і активується при зниженні рівня доступних вуглеводів або АТФ, стимулюючи катаболічні процеси, мобілізацію резервних сполук і автофагію, одночасно пригнічуючи енергозатратні анаболічні процеси. На противагу цьому, TOR є сенсором енергетичного і поживного достатку і активує синтез білків, ріст клітин і проліферацію, забезпечуючи накопичення біомаси. GCN2 виконує функцію сенсора дефіциту амінокислот і обмежує трансляцію білків, дозволяючи клітині економити ресурси і адаптуватися до метаболічного стресу. Таким чином, баланс між активністю SnRK1 і TOR виступає фундаментальним перемикачем між станами росту і виживання.
Окрім білкових сенсорів, важливу інтеграційну роль виконують метаболіти, які одночасно є джерелами енергії і сигнальними молекулами. Цукри і амінокислоти інформують клітину про рівень доступних ресурсів і визначають напрямок метаболічних потоків. Поліаміни, мелатонін і гамма-аміномасляна кислота (GABA) виконують функції регуляторів редокс-стану і стабілізаторів клітинних структур, беручи участь у детоксикації реактивних форм кисню та регуляції експресії генів стресової відповіді. Ці молекули дозволяють клітині швидко адаптувати метаболізм до змін середовища і підтримувати функціональну стабільність.
У цьому контексті хлоропласти виступають ключовими інтеграторами клітинної відповіді, оскільки вони одночасно є головним джерелом енергії і центром генерації сигнальних молекул. У процесі фотосинтезу вони синтезують АТФ і відновлювальні еквіваленти, необхідні для всіх біосинтетичних процесів, але водночас продукують реактивні форми кисню, які виконують роль вторинних месенджерів у сигнальних каскадах. Завдяки цьому хлоропласти забезпечують тісну інтеграцію енергетичного метаболізму і системи імунної відповіді, дозволяючи клітині координувати використання ресурсів залежно від умов.
Мітохондрії є головним центром клітинного виживання, оскільки забезпечують синтез АТФ, підтримання редокс-балансу, регуляцію метаболічної гнучкості та інтеграцію стресових сигналів. Умови стресу часто обмежують фотосинтез, але виживання клітини визначається здатністю мітохондрій підтримувати енергетичний гомеостаз і перебудовувати метаболізм.
Таблиця 7
Молекулярно-клітинні механізми дії антистресових молекул із їхнім впливом на врожайність
| Група / Речовина | Молекулярний механізм | Клітинний ефект | SOURCE–SINK вплив | Фази застосування (пшениця) | Гормональні шляхи | Вплив на врожайність / практичний результат |
| Азотні метаболічні інтегратори: глутамін, аргінін | Донори азоту, підтримка C/N балансу, синтез амінокислот і поліамінів | Підтримка синтезу білків, ремобілізація ресурсів, стабілізація енергетичного статусу клітини | Підвищення sink strength, ефективний перенос сахарози до колоса та зерна | Кущення – прапорцевий лист | TOR/ABA, взаємодія з SnRK1 | Збереження sink strength, ефективний налив зерна, +10–20 % урожаю |
| Осмопротектори: пролін | Осмотичний баланс, стабілізація мембран та білків, захист від дегідратації | Підтримка тургору, стабілізація хлоропластів та органел, захист ферментів | Покращення транспорту асимілянтів, підтримка sink strength | Кущення – початок колосіння | ABA, частково JA | Зменшення втрат урожаю при посусі, покращення наливу і маси зерна |
| Кватернарні амонієві сполуки: гліцин-бетаїн | Мембранна стабілізація, захист фотосистем | Захист PSII, стабілізація тилакоїдів, запобігання дисоціації фотосистем | Підтримка фотосинтетичного потоку → зерно | Кущення – прапорцевий лист | ABA, частково SA | Підтримка маси зерна та крохмалю, +8–15 % врожайності |
| Поліоли / цукрові осмоліти: трегалоза, сахароза, манітол, сорбітол, інозитол | Осмотична регуляція, детоксикація ROS, стабілізація білків | Підтримка тургору, стабілізація ферментів, захист хлоропластів | Покращення транспорту асимілянтів, підтримка sink | Прапорцевий лист – початок наливу | TOR/ABA | Підвищення наливу зерна, стабільність продуктивності, +10–20 % урожайності |
| Антиоксиданти ферментативні: SOD (супероксиддисмутаза), CAT (каталаза), APX (аскорбат-пероксидаза), GPX (глутатіон-пероксидаза) | Каталіз ROS-детоксикації, регенерація редокс-систем | Захист білків, мембран, хлоропластів, підтримка фотосинтезу | Стабільний фотосинтетичний потік → зерно, підтримка sink strength | Кущення – початок колосіння | SA/JA, взаємодія з ABA | Стабільність фотосинтезу, збереження sink strength, +10–18 % урожай |
| Антиоксиданти нефементативні: аскорбат, глутатіон, токофероли, каротиноїди, флавоноїди | Redox-буфер, нейтралізація ROS, регенерація ферментів | Підтримка редокс-гомеостазу, захист генеративних органів | Підтримка фотосинтезу та ефективного наливу | Прапорцевий лист – початок наливу | SA/JA | Підтримка наливу, маси зерна та білка, +8–15 % врожай |
| Сірковмісні попередники антиоксидантів: цистеїн | Попередник глутатіону, детоксикація ROS | Підтримка антиоксидантного захисту, стабілізація ферментів | Підтримка sink strength через стабільний метаболізм | Кущення – прапорцевий лист | TOR/ABA | Підвищення стресостійкості, стабільність продуктивності |
Антистресові речовини рослин виконують критично важливу роль у підтримці гомеостазу клітини та інтегрованої адаптації до абіотичних і біотичних стресів. Їх дія реалізується через кілька взаємопов’язаних механізмів на молекулярному та клітинному рівні. Ферментативні антиоксиданти (SOD – супероксиддисмутаза, CAT – каталаза, APX – аскорбатпероксидаза, GPX – глутатіонпероксидаза) виконують детоксикацію реактивних форм кисню (ROS). SOD каталізує дисмутацію супероксидного аніону в пероксид водню, який далі розщеплюється CAT та пероксидазами, включно з APX та GPX, до води, запобігаючи оксидативному пошкодженню білків, ліпідів та ДНК. Неферментативні антиоксиданти (аскорбат, глутатіон, токофероли, каротиноїди, флавоноїди) діють як редокс-буфери, нейтралізуючи ROS без витрат енергії, відновлюючи окислені ферментативні системи, підтримуючи редокс-гомеостаз, який критично важливий для стабільності фотосинтезу та розвитку генеративних органів.
Осмопротектори, включаючи пролін, гліцин-бетаїн та поліоли (трегалоза, манітол, сорбітол, інозитол), стабілізують клітинні структури та білкові комплекси під час осмотичного та водного стресу. Вони підвищують осмотичний потенціал клітини, підтримують тургор, запобігають дегідратації хлоропластів та інших органел, а також стабілізують мембранні білки та ферментативні комплекси. Гліцин-бетаїн, як кватернарна амонієва сполука, особливо ефективно захищає фотосистеми, запобігаючи дисоціації комплексів PSII і стабілізуючи тилакоїдні мембрани.
Мелатонін як універсальний антистрес-біостимулятор — активно регулює ROS, гормони, та гомеостаз і вже показує ефективність в різних стресах. Мелатонін (N-ацетил-5-метокситриптамін) — це індольна біологічно активна молекула, похідна триптофану. У рослин його відкрили значно пізніше, ніж у тварин, але зараз він визнаний універсальним регулятором стресостійкості та метаболічного гомеостазу. Сьогодні у сільському господарстві вже є комерційні форми мелатоніну, хоча вони ще не так поширені, як класичні стимулятори росту (Plant Melatonin). Триптофан (L‑триптофан) — це незамінна амінокислота, що є попередником багатьох біологічно активних молекул у рослинах і тварин, таких як мелатонін, серотонін, індіол‑3‑оцтова кислота (IAA) — основний ауксин, алкалоїди та інші вторинні метаболіти.
Мелатонін вважається надзвичайно перспективним – він ефективно регулює ROS і гормональний статус у різних стресових умовах. Наприклад, у посушливих дослідах обробка мелатоніном підвищувала врожайність пшениці, ячменю, кукурудзи на 30–100%. Комбіновані формуляції, що поєднують осмопротектанти, антиоксиданти і гормональні модулі, можуть давати синергічний ефект.

Фото 4. Енергетичний колапс клітини.
GABA (γ-аміномасляна кислота) — це стрес-індукований метаболіт, що підтримує клітинний гомеостаз через регуляцію енергетики, редокс-балансу та іонної стабільності. Вона активується за абіотичних і біотичних стресів, підтримує мітохондріальне дихання (через GABA-шунт), знижує накопичення ROS і модулює сигнальні процеси, сприяючи інтегрованій адаптації рослини. При абіотичних стресах (посуха, засолення, спека) GABA сприяє збереженню енергетичного балансу та осмотичної стабільності. При біотичних стресах вона бере участь у сигнальній взаємодії з патогенами та регулює захисні відповіді.
Глутамін та аргінін виступають як донори азоту та C/N балансуючі метаболіти, що підтримують синтез амінокислот, білків, поліамінів і нуклеотидів. Це критично важливо для регуляції Sink strength, ремобілізації резервів та підтримки енергетичного потенціалу клітини під стресом.
Антиоксиданти та осмопротектори взаємодіють із сигналізацією ROS, ABA та TOR/SnRK1, формуючи системні вузли регуляції, які дозволяють рослині адаптивно перемикатися між фазами виживання та росту. Осмопротектори та антиоксиданти знижують оксидативне та осмотичне навантаження, що підтримує транспорт сахарози через SUT і SWEET, зміцнює sink strength зерна та забезпечує стабільне накопичення крохмалю і білка.
У польових умовах застосування цих речовин у критичні фази росту (кущення, прапорцевий лист, налив зерна) дозволяє підвищити стабільність врожайності на 10–25 %, особливо при посусі, високих температурах або солонцевих ґрунтах. Інтеграція антиоксидантної та осмопротекторної підтримки з регуляторними гормональними програмами (цитокініни, ABA) дозволяє керовано формувати SOURCE–SINK баланс, зберігаючи активність листя та забезпечуючи ефективний перенос асимілятів у зерно.
Сила акцепторних органів (sink strength) визначається активністю транспортерів сахарози SUT і SWEET та ферменту інвертаза клітинної стінки (cell wall invertase), які забезпечують транспорт і необоротне використання сахарози в репродуктивних тканинах. Cell wall invertase відіграє ключову роль у формуванні градієнта концентрації, що забезпечує безперервний потік асимілятів у зерно. Пригнічення активності цих білків під дією стресу є головною причиною зниження врожаю навіть за відносно збереженого фотосинтезу.
Ефективні антистресові технології реалізують свій позитивний вплив через підтримання функціонального SOURCE–SINK балансу. Це включає збереження фотосинтетичної активності SOURCE-органів, підтримання транспорту асимілятів через флоему і забезпечення нормального розвитку SINK-органів, зокрема зерна. Саме безперервний потік асимілятів визначає енергетичний статус рослини і її здатність підтримувати ріст і формування врожаю в умовах стресу.
Ключовим механізмом відновлення продуктивності при стресі є активація SOURCE–SINK взаємодії через стимуляцію транспортерів сахарози SUT і SWEET та ферменту cell wall invertase. Найефективнішими регуляторами цих процесів є цитокініни, калій, бор, амінокислота глутамін, яка є донором азоту, метаболічним інтегратором C/N балансу, амінокислоти-осмопротектори (пролін, гліцин-бетаїн), антиоксиданти (супероксиддисмутаза (SOD), каталаза (CAT), пероксидази (APX, GPX)), неферментативні антиоксиданти (аскорбінова кислота (вітамін C), глутатіон, токофероли (вітамін E), каротиноїди, флавоноїди), розчинні цукри (трегалоза-6-фосфат (T6P), глюкоза, сахароза, манітол), поліоли (сорбітол, інозитол), які безпосередньо підвищують силу акцептора, транспорт асимілятів і налив зерна.
У запропонованій системній моделі регуляції продуктивності та стресостійкості генетичний блок адаптації є обов’язковим рівнем інтеграції, оскільки саме він забезпечує довготривалу перебудову метаболічних і фізіологічних процесів. Якщо TOR ↔ SnRK1 та гормональні сигнали визначають оперативне перемикання між ростом і захистом, то транскрипційні фактори й захисні білки формують стійку програму відповіді. Ключову роль у цьому відіграють родини генів DREB, NAC та HSP.
Гени родини DREB (Dehydration-Responsive Element Binding) кодують транскрипційні фактори, що розпізнають специфічні DRE/CRT-елементи у промоторах стрес-індукованих генів. Вони активуються за умов водного дефіциту, низьких температур і осмотичного стресу та ініціюють експресію широкого спектра захисних генів — від синтезу осмопротекторів (пролін, цукри) до ферментів антиоксидантної системи. Функціонально DREB забезпечують швидке перепрограмування клітинного метаболізму в умовах падіння енергетичного статусу. У контексті моделі TOR ↔ SnRK1 активація DREB тісно пов’язана зі зниженням активності TOR і переходом у SnRK1-домінантний стан. Це сприяє збереженню клітинної гомеостазії, але водночас може тимчасово обмежувати ростові процеси та інтенсивність SOURCE→SINK транспорту. Для агрономії це означає, що високий потенціал DREB-опосередкованої відповіді підвищує стійкість до посухи та холоду, але потребує технологічного супроводу для мінімізації втрат урожайності.
Транскрипційні фактори родини NAC (NAM, ATAF, CUC) мають ширший функціональний спектр. Вони беруть участь у регуляції старіння листків, ремобілізації азоту та вуглецю, формуванні кореневої системи та адаптації до посухи й солонцевих умов. У генеративних фазах розвитку NAC-фактори контролюють перерозподіл ресурсів із SOURCE-органів до SINK-органів, визначаючи ефективність наливу зерна. За умов стресу окремі представники родини NAC активують гени антиоксидантного захисту та білки, пов’язані з детоксикацією реактивних форм кисню. В енергетичному вимірі їхня активація часто асоціюється з переходом системи до консервативного режиму, але водночас вони можуть забезпечувати більш ефективну ремобілізацію ресурсів, що частково компенсує зниження фотосинтетичної активності. Таким чином, NAC виступають містком між стресостійкістю та врожайністю, особливо в умовах обмеженого водного забезпечення.
Гени HSP (Heat Shock Proteins) кодують теплові шокові білки — молекулярні шаперони, що стабілізують третинну та четвертинну структуру білків під час температурного або окисного стресу. Вони запобігають денатурації ферментів фотосинтезу, дихального ланцюга та інших ключових метаболічних комплексів. HSP забезпечують підтримання функціональної активності клітин навіть за умов теплових хвиль, що особливо критично в період цвітіння та наливу зерна. З позиції метаболічного ландшафту HSP не змінюють напрямок переходу між TOR- і SnRK1-станами, але зменшують ймовірність різкого критичного переходу, стабілізуючи ферментативні системи та знижуючи рівень «метаболічного шуму». Це дозволяє культурі довше утримуватися в продуктивному режимі навіть за короткочасних температурних піків.
Інтеграція генетичного блоку DREB–NAC–HSP у модель TOR ↔ SnRK1 доповнює її часовим виміром і визначає глибину й тривалість адаптаційної відповіді. Якщо гормональні сигнали та енергетичні сенсори відповідають за швидке перемикання станів, то транскрипційні фактори та шаперони забезпечують стабілізацію нового стану й формують адаптивну пам’ять. Для практичної агрономії це означає, що вибір гібридів із підвищеною експресією або ефективною регуляцією цих генів може суттєво підвищити стабільність урожайності в умовах кліматичної нестабільності. Поєднання генетичного потенціалу (DREB, NAC, HSP), збалансованої агротехнології та управління енергетичним статусом рослини дозволяє сформувати систему, у якій стресостійкість не протиставляється врожайності, а стає інструментом її стабілізації.
Фактично клітина функціонує як складна інтеграційна захисна система, яка постійно оцінює інтенсивність і тривалість стресового сигналу та приймає рішення щодо подальшої стратегії — адаптації, відновлення або програмованої загибелі. Центральним механізмом цієї адаптації є автофагія — еволюційно консервативний процес деградації і рециклінгу клітинних компонентів. Автофагія забезпечує видалення пошкоджених білків, ліпідів і дисфункціональних органел, включаючи хлоропласти і мітохондрії, тим самим запобігаючи накопиченню токсичних продуктів. Водночас продукти деградації повторно використовуються як джерело енергії і будівельних блоків, що дозволяє клітині підтримувати метаболізм навіть у критичних умовах. Особливе значення автофагія має під час гіперчутливої відповіді, де вона може обмежувати поширення пошкодження або забезпечувати контрольовану утилізацію клітинних структур.
Паралельно функціонує система підтримання протеостазу, яка включає убіквітин-протеасомну систему, ER-асоційовану деградацію і відповідь на неправильно згорнуті білки. Ці механізми забезпечують постійний контроль якості протеому, розпізнаючи і видаляючи пошкоджені або нефункціональні білки. Це дозволяє клітині швидко перебудовувати свій функціональний стан, адаптуючись до нових умов.
Важливу координаційну роль відіграє цитоскелет, сформований актиновими філаментами і мікротрубочками, який забезпечує транспорт органел, сигнальних комплексів і автофагосом. Завдяки цитоскелету клітина може швидко перерозподіляти ресурси і координувати захисні процеси у просторі і часі. Перебудова цитоскелету є важливим елементом адаптації, оскільки вона впливає на стабільність клітинних структур і ефективність внутрішньоклітинної комунікації.
Ключовим елементом виживання є також антиоксидантна система, яка підтримує редокс-гомеостаз клітини. Ферменти, такі як супероксиддисмутаза, каталаза, аскорбатпероксидаза і глутатіонпероксидаза, разом із низькомолекулярними антиоксидантами нейтралізують надлишкові реактивні форми кисню, запобігаючи пошкодженню клітинних компонентів. Контроль рівня ROS визначає, чи клітина зможе адаптуватися, чи перейде до програмованої загибелі.
Додаткову стабілізацію забезпечують осмо-іонні і метаболічні регулятори, включаючи іони калію і хлору, а також пролін, мелатонін і GABA. Ці сполуки підтримують осмотичний баланс, стабілізують білки і мембрани та координують метаболічну адаптацію до стресу.
Уся ця багаторівнева захисна програма клітини, що включає внутрішньоклітинні механізми координації швидких і локальних відповідей на стрес, окремі елементи якої були розглянуті вище у спрощеному вигляді — від молекулярних сенсорів і сигнальних каскадів до систем підтримання енергетичного та структурного гомеостазу, — у своїй сукупності формує цілісну інтегровану систему імунітету рослини до патогенів і стресів. При цьому захисна відповідь не обмежується межами окремої клітини, а поширюється на рівень тканин і всього організму. На рівні цілісної рослини та її взаємодії з довкіллям імунітет набуває системного характеру, в межах якого локальні клітинні сигнали трансформуються у координовані організмові реакції. Така трансформація забезпечується механізмами системної інтеграції, що узгоджують функціонування різних органів, тканин і мікрооточення, формуючи єдину адаптивну відповідь на дію стресових факторів.
Імунна мережа функціонує не у вакуумі, а в екологічному контексті. Мікробіом ризосфери та філосфери є фундаментом для певного типу імунітету ISR від некротрофних патогнів і стресів. Вторинні метаболіти (фіталексини, фенілпропаноїди, терпенові леткі) не тільки діють проти патогенів, але й впливають на сусідні рослини й мікробні спільноти. Індукований системний захист (ISR, Induced Systemic Resistance) активується коменсальними або симбіотичними мікроорганізмами, що взаємодіють із кореневою системою. ISR підвищує готовність рослини до атак патогенів, стимулюючи гормональні шляхи JA/ET та мобілізацію ROS/NO у віддалених тканинах.
Мікробіом як модулюючий фактор екології виконує ключову роль у регуляції системного імунітету. Симбіотичні бактерії та гриби формують захисний “бар’єр” проти патогенів, стимулюють ISR, покращують поживну та водну забезпеченість рослини, а також впливають на гормональні та метаболічні шляхи. Взаємодія з мікробіомом дозволяє рослині адаптуватися до мінливих умов середовища, підвищує крос-толерантність і сприяє довготривалому виживанню організму на рівні екосистеми.
На рівні організму ця захисна система функціонує не як набір ізольованих реакцій, а як єдина інтегрована регуляторна мережа, яку ми визначаємо як інтегративну модель імунітету рослин до патогенів і стресів SA–JA/ET–ABA–TOR. Її ключова особливість полягає у здатності безперервно оцінювати інтенсивність, тривалість і природу стресового сигналу та на основі цього приймати стратегічне рішення щодо подальшої долі клітини і рослини в цілому — переходу до адаптації, активації відновлення або, у разі критичних пошкоджень, запуску програмованої клітинної загибелі як захисного механізму.

Фото 5. Від росту до виживання.
Функціонально ця система являє собою мультистабільну сигнально-енергетично-метаболічну мережу, яка сформувалася в процесі еволюції під постійним тиском біотичних і абіотичних факторів середовища. На організмовому рівні її можна описати як динамічний «ландшафт станів», у межах якого клітина і тканини здатні переходити між різними функціональними режимами залежно від типу і сили стресу. У цьому ландшафті SA-домінантний стан забезпечує ефективний імунний захист проти біотрофних патогенів шляхом активації системної набутої резистентності, синтезу захисних білків і локалізації інфекції. Натомість JA/ET-домінантний режим спеціалізований на протидії некротрофним патогенам, механічним пошкодженням і травматичному стресу, координуючи синтез вторинних метаболітів, посилення бар’єрних структур і репаративні процеси. У свою чергу, ABA-домінантний стан відіграє ключову роль у забезпеченні адаптації до абіотичних факторів, таких як посуха, засолення, температурні екстремуми та порушення водного балансу, спрямовуючи метаболізм на збереження ресурсів і стабілізацію клітинних структур.
Окреме, принципово важливе місце в цій інтегративній моделі займає TOR-домінантний стан, який визначає ростовий режим функціонування рослини і формує основний внесок у майбутню врожайність. Якщо SA-, JA/ET- та ABA-домінантні стани спрямовані передусім на захист і виживання, то TOR-стан забезпечує реалізацію продуктивного потенціалу, контролюючи процеси біосинтезу, формування біомаси і генеративних органів. Саме тому цей стан є ключовою фізіологічною передумовою формування високого врожаю.
TOR-стан є функціонально антагоністичним до більшості захисних програм, оскільки останні потребують значних енергетичних витрат і супроводжуються пригніченням ростових процесів. Активація імунітету, синтез захисних білків, антиоксидантів і вторинних метаболітів, а також процеси автофагії і репарації неминуче відволікають ресурси від синтезу структурної біомаси. У противагу цьому, TOR функціонує як центральний сенсор енергетичного і метаболічного достатку, і його активація сигналізує клітині про наявність сприятливих умов для росту і розвитку.
Ростовий TOR-домінантний стан активується за відсутності стресових факторів або після їх ефективної нейтралізації та відновлення енергетичного балансу клітини. У цьому режимі відбувається системне переключення метаболізму на анаболічні процеси: різко зростає інтенсивність синтезу білків, нуклеїнових кислот, ліпідів і компонентів клітинної стінки, активується робота рибосом і прискорюється клітинний цикл. Це забезпечує інтенсивний поділ і диференціацію клітин, ріст меристем, формування листкової поверхні, розвиток кореневої системи і генеративних органів, які безпосередньо визначають рівень майбутньої врожайності.
Гормональна регуляція цього стану характеризується домінуванням фітогормонів росту, зокрема ауксинів (IAA), гіберелінів (GA), брассиностероїдів (BR) і цитокінінів (CK). Ауксини стимулюють розтягування клітин і формування провідних тканин, гібереліни активують поділ клітин і ріст стебла, брассиностероїди підсилюють фотосинтетичну активність і синтез біомаси, а цитокініни стимулюють поділ клітин і затримують старіння тканин. Сукупна дія цих гормонів створює фізіологічні умови для максимальної реалізації ростового потенціалу.
Одночасно з цим у TOR-домінантному стані відбувається тимчасове зниження активності захисних механізмів. Це не означає втрату імунітету, а є стратегічною оптимізацією використання ресурсів, при якій енергія і метаболіти спрямовуються передусім на формування продуктивних структур. Таким чином, рослина переходить із режиму виживання у режим продуктивного функціонування, що є критичною умовою формування високої врожайності.
У фізіологічному і агрономічному контексті це означає, що максимальна врожайність формується не в умовах постійної активації захисних програм, а внаслідок своєчасного і ефективного переключення рослини у TOR-домінантний ростовий стан після завершення або нейтралізації стресу. Саме тривалість і повнота перебування рослини в цьому стані визначають інтенсивність накопичення біомаси, ефективність фотосинтезу, формування репродуктивних органів і, як наслідок, кінцевий рівень врожаю. У цьому контексті TOR виступає не лише регулятором клітинного росту, а центральним інтегратором продуктивності, який визначає, наскільки повно рослина зможе реалізувати свій генетично закладений потенціал урожайності.
Важливо, що переходи між цими функціональними станами не є випадковими, а визначаються скоординованою роботою фундаментальних внутрішньоклітинних інтеграторів, включаючи енергетичні сенсори SnRK1 і TOR, систему автофагії, антиоксидантний захист, механізми протеостазу та динамічну організацію цитоскелету. Саме ці механізми забезпечують здатність клітини гнучко перерозподіляти енергетичні і метаболічні ресурси, видаляти пошкоджені компоненти, стабілізувати функціонування ключових органел і координувати просторово-часову організацію відповіді на стрес.
У результаті формується високоефективна адаптивна система, яка дозволяє рослині не лише реагувати на окремі стресові фактори, а й інтегрувати їх у єдину відповідь, оптимізуючи баланс між процесами росту, захисту і відновлення. Саме завдяки функціонуванню цієї інтегративної мережі рослина здатна підтримувати внутрішній гомеостаз, зберігати життєздатність клітин і забезпечувати довготривалу продуктивність навіть у мінливих і несприятливих умовах середовища. У цьому контексті імунітет рослин постає не лише як система захисту, а як фундаментальний механізм управління життєвим циклом, який визначає здатність рослини реалізувати свій генетичний потенціал продуктивності.
Як керувати балансом «ріст – захист» через живлення?
Наші ключові наукові дослідження останніх років переконливо демонструють, що фундаментальною основою формування врожайності є баланс між двома центральними регуляторними системами клітини — TOR і SnRK1, які виконують роль головних метаболічних перемикачів між режимами росту і виживання від стресів.
Ці режими рослини антагоністичні. Рослина не може одночасно активно рости й повноцінно захищатися. Неправильне втручання — наприклад, стимуляція TOR під час стресу або імунної атаки — призводить до втрат урожайності на 10–45%, навіть за повного забезпечення добривами й пестицидами.
Комплекс TOR (Target of Rapamycin) визначає потенціал продуктивності рослини, оскільки саме він контролює інтенсивність клітинного поділу, швидкість росту, формування біомаси та закладання генеративних органів, включаючи кількість і масу зерна. Коли TOR перебуває в активному стані, рослина працює в режимі максимального синтезу, ефективно використовує ресурси для нарощування вегетативної маси та формування врожаю. Практичні й експериментальні дані показують, що застосування факторів, які активують TOR-сигналінг, зокрема доступного нітратного азоту, амінокислот, вуглеводів і цитокінінів, дозволяє підвищити врожайність у середньому на 15–40 %, що пов’язано з посиленням біосинтетичних процесів і більш повною реалізацією генетичного потенціалу культури.
Водночас система SnRK1 виконує принципово іншу, але не менш важливу функцію. Вона є ключовим сенсором енергетичного дефіциту і активується в умовах стресу, коли порушується фотосинтез, обмежується доступ води або виникають температурні чи хімічні пошкодження. Основна роль SnRK1 полягає у збереженні життєздатності клітин шляхом пригнічення енерговитратних процесів росту і одночасної активації захисних механізмів, включаючи антиоксидантний захист, стабілізацію мембран і мобілізацію резервних ресурсів. Завдяки цьому рослина здатна пережити несприятливий період з мінімальними втратами. За результатами польових і фізіологічних досліджень, ефективна активація SnRK1-залежних механізмів дозволяє зберегти від 10 до 30 % врожаю, який в іншому випадку був би втрачений через загибель клітин і незворотне пошкодження фотосинтетичного апарату.
Проте найбільш важливим і практично значущим нашим відкриттям є те, що максимальна врожайність формується не при постійній активації лише системи росту або лише системи захисту, а саме при їх правильному і своєчасному перемиканні. Рослина повинна спочатку активувати SnRK1 для переживання стресу і стабілізації клітинних структур, а після відновлення сприятливих умов — переключитися на домінування TOR, що забезпечує інтенсивний ріст і формування врожаю. Саме така динамічна рівновага між режимом збереження і режимом продуктивності дозволяє досягти максимального результату. Практичні дані свідчать, що оптимальне управління балансом SnRK1–TOR забезпечує сумарне підвищення врожайності на рівні 20–50 %, що є найвищим показником серед усіх сучасних фізіологічних підходів до управління продуктивністю рослин.
Таким чином, критично важливий практичний висновок полягає в тому, що максимальна врожайність визначається не максимальною швидкістю росту як такою, а здатністю рослини своєчасно переключатися між програмами виживання і продуктивності. Ефективне управління цим перемиканням — від активації SnRK1 у період стресу до домінування TOR у фазі відновлення — є фундаментальною основою сучасних технологій стабільного і високопродуктивного рослинництва.
В інтегративній моделі імунітету чітко демонструється, що багато помилок у агротехнологіях виникає не через вибір препаратів чи норми внесення, а через передчасну або невчасну активацію TOR-сигналізації. При цьому TOR не є ані «шкідливим», ані «корисним» сам по собі: його функція завжди визначається контекстом і поточним фізіологічним станом рослини. Це усвідомлення відкриває нову парадигму в агрономії: ключовим чинником ефективності є не самі пестициди чи добрива, а правильний час, коли рослині дозволяють знову перейти в TOR-залежний стан росту та продуктивності. Це одна з ключових ідей в інтегративній моделі імунітету рослин.
Таблиця 8
Практична інтерпретація для агрономії
| Період | Активний шлях | Що застосовувати | Ефект |
| Перед стресом | SnRK1 | К, Si, Mg | Захист |
| Під час стресу | SnRK1 | антистресанти | Виживання |
| Після стресу | TOR | амінокислоти + NO₃ | Відновлення |
| Період росту | TOR | N, Mg | Максимальний урожай |
Розроблена нами на підставі інтегративної моделі імунітету рослин від патогенів і стресів SA–JA/ET–ABA–TOR практична концепція «Імунно-метаболічного управління культурою» пропонує працювати не з окремими симптомами, а з регуляторними режимами рослини. Традиційні біостимулятори формувалися у рамках підходу, який розглядає рослину як пасивний об’єкт: достатньо збільшити надходження «ресурсів», щоб активувати ріст. У цій парадигмі амінокислоти, гумінові сполуки чи мікроелементи нібито автоматично конвертуються у біомасу та врожай. Проте сучасна фізіологія рослин демонструє протилежне: за стресових умов така схема не працює. Рослина свідомо гальмує ростові програми, пригнічуючи TOR-сигналізацію, і переходить у захисні режими виживання, керовані SA, JA/ET та ABA.
Регуляторні технології базуються на принципово іншому підході. Вони розглядають рослину як активну, саморегульовану систему з чітко визначеними функціональними станами. Мета таких технологій не у примусовому рості за будь-яких умов, а в управлінні переходами між фазами «стрес → стабілізація → відновлення». Ключова роль відводиться фазі стабілізації, де домінують гормональні каскади ABA, SA та JA. Лише після відновлення метаболічної рівноваги можливо повторне увімкнення TOR-залежних механізмів росту та продуктивності.
Фактично класичні біостимулятори «підштовхують» рослину до росту незалежно від її фізіологічного стану. Регуляторні ж технології діють точніше: спершу забезпечують стабілізацію та баланс, а вже потім дозволяють ефективно відновити ріст і реалізувати врожайний потенціал. Цю різницю можна порівняти з людським організмом: класичні біостимулятори – це як змушувати людину з лихоманкою та зневодненням бігти марафон, тоді як регуляторні технології спершу «лікують» — відновлюють баланс і стабільність — і лише після цього повертають організм до високої продуктивності.
Стрес у рослин слід розглядати не як зовнішню подію — посуху, патоген або температурний удар — а як специфічний внутрішній режим регуляції. Цей підхід означає парадигмальний зсув: стрес перестає бути синонімом фактору впливу і визначається як гормонально-метаболічний стан, у якому перебудовується сигнальна, енергетична та ростова архітектура рослини. Завдяки цьому стає можливим класифікувати стресові стани за домінуючими сигнальними шляхами — SA, JA, ABA або TOR — замість орієнтації на зовнішні фактори.
Інтегративна модель імунітету рослин показує: більшість антистресантів насправді не «знімають» стрес, а лише перемикають рослину в інший фізіологічний режим, який може бути невідповідним її реальному стану. Тому сам термін «антистресанти» як клас продуктів є метаболічно некоректним, оскільки створює ілюзію універсальної дії там, де відбувається тонке втручання у регуляторні системи рослини. Цей підхід дозволяє сформувати нову, біологічно обґрунтовану класифікацію продуктів — не за декларативним «ефектом», а за регуляторним режимом, який вони активують: режими виживання (ABA), імунного захисту (SA або JA/ET) або фаза росту і відновлення (TOR).
В результаті наукова і практична агрономія переходить від інтуїтивного застосування «універсальних» продуктів до свідомого управління фізіологічними станами рослини. Кожне втручання має чітку мету, визначену фазу та внутрішню логіку, що формує нову регуляторну парадигму, де агротехнології працюють у синхронії з біологією рослини.
Замість застосування універсальних «антистресантів» концепція пропонує двофазний підхід до управління культурою:
- Фаза стабілізації. Активується відповідний імунний або стресовий режим (SA, JA/ET або ABA), який забезпечує збереження клітин, підтримку енергетичного балансу та базової життєздатності рослини.
- Фаза відновлення. Лише після нейтралізації стресових факторів відбувається контрольована активація TOR, що повертає рослину до росту, фотосинтетичної активності та повноцінної реалізації врожайного потенціалу.
Такий підхід дозволяє відійти від календарних схем і перейти до фізіологічно обґрунтованого управління, де кожне втручання синхронізоване з фактичним імунно-метаболічним станом культури.
Баланс між ростом і захистом у рослин є фундаментальним фізіологічним процесом, який безпосередньо визначає їх продуктивність в умовах стресу. Оскільки будь-яка стресова відповідь потребує значних енергетичних і метаболічних витрат, рослина змушена постійно перерозподіляти ресурси між процесами росту та механізмами виживання. Центральну роль у цьому регуляторному механізмі відіграють дві взаємопов’язані сигнальні системи — TOR (Target of Rapamycin) і SnRK1 (SNF1-related kinase 1), які функціонують як молекулярні сенсори енергетичного і метаболічного стану клітини. TOR активується в умовах достатнього забезпечення енергією і поживними речовинами та стимулює анаболічні процеси, включаючи синтез білка, поділ клітин і активний ріст. Навпаки, SnRK1 активується при енергетичному дефіциті або стресі та переводить метаболізм у режим економії, пригнічуючи ріст і активуючи захисні механізми.
З практичної точки зору, управління цим балансом можливе через цілеспрямоване застосування елементів живлення і біологічно активних сполук. Для стимуляції росту та активації TOR-сигналінгу найбільш ефективними є нітратна форма азоту (NO₃⁻), легкодоступні цукри, амінокислоти та цитокініни. Ці сполуки сигналізують рослині про достатній рівень енергії та ресурсів, що призводить до активації синтезу білка, відновлення фотосинтетичної активності та формування нових клітин. Тому їх застосування є особливо доцільним після дії стресових факторів, таких як гербіцидний вплив, посуха або низькі температури, коли необхідно якнайшвидше відновити нормальний метаболізм і ріст. У таких умовах оптимальним практичним рішенням є використання амінокислот у поєднанні з нітратним азотом, що дозволяє швидко активувати TOR-залежні процеси і прискорити регенерацію рослин.
Водночас для підвищення стійкості рослин до стресових факторів і активації захисних механізмів, пов’язаних із SnRK1, ефективними є інші елементи і метаболіти, зокрема калій, кремній, магній, поліаміни та мелатонін. Калій відіграє ключову роль у підтриманні осмотичного балансу і регуляції роботи продихів, що особливо важливо під час посухи або температурного стресу. Кремній зміцнює клітинні стінки та підвищує механічну і фізіологічну стійкість тканин. Магній забезпечує стабільність фотосинтетичного апарату і підтримує синтез АТФ, тоді як поліаміни і мелатонін виконують функції потужних антиоксидантів, знижуючи рівень реактивних форм кисню і захищаючи клітинні структури від пошкодження. Найбільш ефективним є їх застосування до настання стресу або на ранніх етапах його дії, коли ще можливо активувати захисні системи і мінімізувати потенційні пошкодження. Наприклад, перед прогнозованим заморозком практично обґрунтованим заходом є позакореневе внесення калію в поєднанні з кремнієм, що дозволяє значно підвищити стійкість рослин до низьких температур.
Таким чином, регуляція балансу між ростом і захистом через систему живлення є ефективним інструментом управління фізіологічним станом рослин. Застосування сполук, що активують TOR, є доцільним у період відновлення після стресу для стимуляції росту і продуктивності, тоді як використання елементів, що активують SnRK1, дозволяє підвищити стійкість рослин і підготувати їх до несприятливих умов. Раціональне поєднання цих підходів забезпечує оптимальний метаболічний баланс, збереження фотосинтетичної активності і максимальну реалізацію продуктивного потенціалу культури навіть в умовах стресу.
Таблиця 9
Управління балансом «ріст – захист» через живлення рослин
| Фізіологічний напрям | Сигнальний шлях | Елементи / речовини | Основний фізіологічний ефект | Коли застосовувати | Практична мета | Практичний приклад |
| Стимуляція росту | TOR (Target of Rapamycin) | Нітратний азот (NO₃⁻) | Активує синтез білків, поділ клітин, ріст біомаси | Після стресу | Швидке відновлення росту | Після гербіцидного стресу — внесення нітратного азоту |
| TOR | Цукри | Джерело енергії та сигнальні молекули росту | Після стресу | Відновлення енергетичного балансу | Позакореневе підживлення у фазу відновлення | |
| TOR | Амінокислоти | Стимулюють синтез білків, відновлення метаболізму | Після стресу | Прискорення регенерації тканин | Після посухи або гербіциду — амінокислоти 1–2 л/га | |
| TOR | Цитокініни | Стимулюють поділ клітин, відновлення росту | Фаза активного відновлення | Запуск нового росту | Відновлення після холодового стресу | |
| Підвищення стійкості | SnRK1 (SNF1-related kinase 1) | Калій (K) | Регулює осмотичний баланс, зменшує втрати води | Перед і під час стресу | Підвищення посухо- і холодостійкості | Перед заморозком — калій 1–2 кг/га |
| SnRK1 | Кремній (Si) | Посилює механічний і антиоксидантний захист | Перед стресом | Підвищення загальної стійкості | Перед посухою або температурним стресом | |
| SnRK1 | Магній (Mg) | Стабілізує фотосинтез і виробництво енергії | Перед і під час стресу | Захист фотосинтетичного апарату | MgSO₄ перед гербіцидом | |
| SnRK1 | Поліаміни | Захищають клітинні структури, стабілізують мембрани | Перед стресом | Підвищення клітинної стійкості | Антистресові композиції | |
| SnRK1 | Мелатонін | Потужний антиоксидант і регулятор стресу | Перед і під час стресу | Зниження оксидативного пошкодження | Антистресові біостимулятори |
Чому це критично важливо саме зараз?
Сучасні кліматичні тренди демонструють, що стресові періоди для культур стають довшими, екстремальні умови повторюються протягом одного сезону, а фаза повного відновлення рослин часто відсутня. У таких умовах здатність керувати стресостійкістю стає визначальним фактором конкурентоспроможності агробізнесу. Абіотичні стреси здатні знижувати врожайність більше ніж на 60%. Тому ринок антистресових технологій зростає рекордними темпами. Однак без науково обґрунтованої логіки ці інвестиції часто не дають очікуваного результату.
Більше ніж концепція — нова філософія агрономії.
«Імунно-метаболічне управління культурою» — це не просто окремий препарат і не чергова агрономічна рекомендація. Це принципово новий підхід до роботи з рослиною, що розглядає її як активну, саморегульовану біологічну систему, здатну перемикатися між режимами росту, захисту та виживання.
Цей підхід:
- пояснює, чому технології, що працювали минулого року, можуть не давати ефекту цього сезону;
- забезпечує агронома чіткою логікою прийняття рішень у складних мультистресових умовах;
- відкриває шлях до створення антистресових технологій нового покоління, заснованих на механізмах, а не на емпіричних обіцянках.
Фактично йдеться про перехід від реактивного захисту до проактивного управління продуктивністю. Саме ті компанії та господарства, які першими інтегрують цю логіку у свої технології, отримають стратегічну перевагу в агровиробництві найближчих років. Настав час перейти на новий рівень — Імунно-метаболічного управління культурою!
Хаблак Сергій, д.б.н., агроном-експерт, агроаудитор, 066-44-266-08
Читайте нас у TelegramПов’язані теми:










Sumitomo Chemical подала заявки на реєстрацію нового фунгіциду bifemetstrobin у Японії, США та Бразилії
13.07.2026, 11:27
ЗЗР та насіння

Італійські дослідники підтвердили ефективність дріжджового біопрепарату проти несправжньої борошнистої роси винограду
13.07.2026, 09:07
ЗЗР та насіння

Дослідники виявили перспективний біоагент проти фузаріозного в’янення бананів
12.07.2026, 23:41
ЗЗР та насіння

При купівлі вершкового масла приділяйте увагу інформації на маркуванні
12.07.2026, 10:49
Новини