
ІДЕАЛЬНОЇ ФОРМИ МІКРОДОБРИВ НЕ ІСНУЄ
16.05.2026

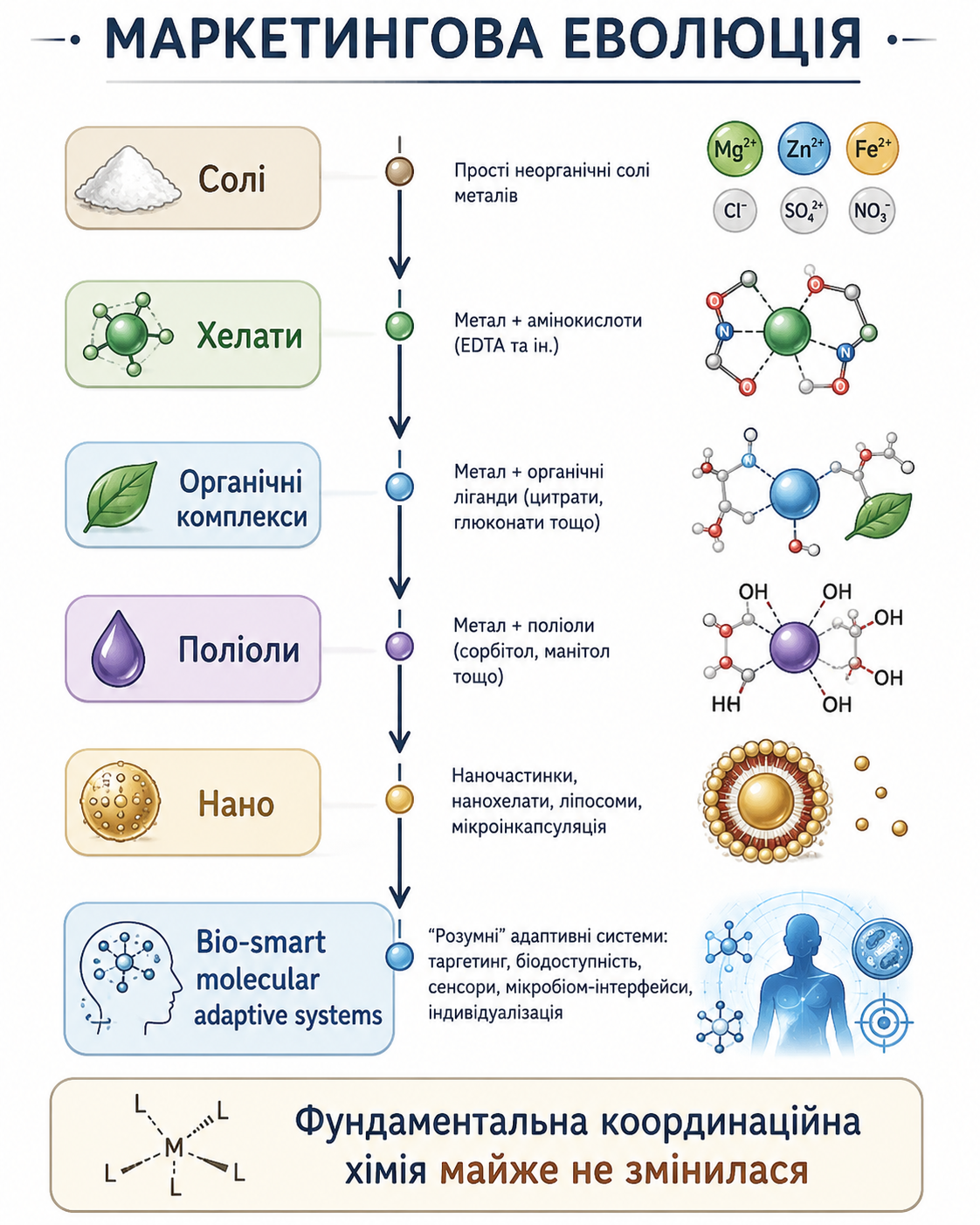
Мал. 1. Маркетингова еволюція технологій доставки мікроелементів. Концептуальна схема еволюції маркетингових підходів до форм мікроелементів — від неорганічних солей і хелатів до наноформ та bio-smart molecular adaptive systems. Попри ускладнення технологічних концепцій, фундаментальні принципи координаційної хімії комплексоутворення залишаються переважно незмінними. У мікроживленні не існує ідеальної форми — існують лише різні способи балансування неминучих компромісів. У реальній агрохімії фермер найчастіше обирає не “кращу” форму мікроелемента, а іншу конфігурацію того самого фізико-хімічного компромісу. У більшості випадків нова форма мікроелемента не усуває фундаментальних обмежень системи, а лише змінює співвідношення між її перевагами та слабкими місцями. У практиці мікроживлення мова найчастіше йде не про “кращу” форму, а про альтернативний компроміс між фізико-хімічними обмеженнями системи.
ВСТУП.
У сучасному агробізнесі мікроелементи майже ніколи не продаються як проста хімічна функція — як іон цинку, марганцю, бору чи міді, необхідний для ферментативних процесів рослини. Натомість споживачу продається не стільки речовина, скільки інтерпретація цієї речовини, тобто “історія про технологію”. Саме тому на етикетках і в презентаціях з’являються терміни на кшталт “нано-формула”, “біоактивний комплекс”, “органічно зв’язаний мікроелемент”, “інтелектуальний хелат” або “технологія нового покоління”, які створюють відчуття принципово нового рівня науки. Однак якщо прибрати термінологічний шар і розглянути хімічну основу таких продуктів, дуже часто виявляється, що мова йде про добре відомі сполуки і класичні принципи координаційної хімії, які використовуються десятки років: хелати на основі EDTA, DTPA або EDDHA, прості солі мікроелементів, комплекси з органічними кислотами, поліолами або амінними похідними.
Реальна різниця між цими продуктами часто полягає не в появі принципово нових механізмів засвоєння рослиною, а в зміні балансу фізико-хімічних властивостей: стабільності в розчині, стійкості до жорсткої води, швидкості дисоціації та сумісності з іншими компонентами бакових сумішей. Але цей баланс у маркетинговій комунікації майже ніколи не пояснюється, оскільки він складний, багатофакторний і не дає простого меседжу. Натомість складність замінюється ідеєю “технологічного прориву”, де будь-яка модифікація молекули подається як якісний стрибок у біодоступності або ефективності.
Ключова проблема полягає в тому, що в агрохімії не існує універсально “кращої” форми мікроелемента, яка одночасно максимально стабільна в баку, максимально швидко проникає через кутикулу і повністю контролюється всередині рослини. Кожна хімічна форма — це компроміс між цими параметрами. Наприклад, більш стабільні хелати краще захищають іон у розчині, але можуть повільніше віддавати його в зоні поглинання; менш стабільні комплекси швидше дисоціюють, але гірше витримують умови жорсткої води або взаємодію з антагоністами. Таким чином, різні формуляції вирішують не одну й ту саму задачу, а різні її частини.
Попри це, у маркетинговій площині ці відмінності часто стираються, і форму подають як самостійний фактор ефективності, ніби “нано” або “органічний комплекс” автоматично означає кращу фізіологічну відповідь рослини. У результаті створюється ілюзія лінійного прогресу, де нова назва дорівнює новому рівню ефективності, хоча насправді йдеться про варіації однієї і тієї ж фундаментальної хімічної системи.
Тому головне питання в реальній агрохімії полягає не в тому, що “працює краще” як абстрактна категорія, а в тому, для яких умов, у якій системі і на якому етапі живлення ця конкретна форма є оптимальною. Саме тут і виникає розрив між маркетинговою риторикою та фізико-хімічною реальністю: індустрія продає ідею універсальної інновації, тоді як наука описує систему компромісів, залежностей і обмежень, які не можна обійти лише зміною назви або формулювання (табл. 1).
Таблиця 1
Порівняльна оцінка форм мікроелементів у системі листкового живлення: функції, обмеження та критичні слабкі місця
| Форма | Основна хімічна роль | Сильна сторона | Слабке місце | Де найчастіше втрачається ефективність | Найбільше обмеження в системі листкового живлення | Типове маркетингове перебільшення |
| Неорганічні солі (сульфати, нітрати) | Джерело вільного іона | Дешеві, швидке вивільнення | Осадження, фітотоксичність, низька стабільність | Бакова суміш, кристалізація після висихання | Низька стабільність у жорсткій воді | «Швидкодіюча форма» |
| EDTA / DTPA хелати | Сильна стабілізація металу | Добра сумісність, захист від антагонізму | Повільніше вивільнення іона | Етап біодоступності біля мембрани | Надмірна стабільність комплексу | «Максимальне засвоєння» |
| EDDHA та сильні хелати | Максимальна стабілізація | Стійкість при високому pH | Часто надлишкова стабільність для foliar | Після проникнення | Ускладнене вивільнення іона | «Преміум доступність» |
| Органічні кислоти (цитрат, глюконат, лігносульфонат) | Слабке комплексоутворення | Швидша дисоціація | Менша стабільність | Бакова суміш, жорстка вода | Осадження/реакції з катіонами | «Натуральне швидке живлення» |
| Поліоли (манітол, сорбітол) | Модифікація фізики краплі, слабке комплексування | Покращення змочування, утримання вологи | Не гарантують проникнення | Кутикулярний бар’єр | Відсутність прямого транспорту через мембрани | «Транспортер у клітину» |
| Цукрові комплекси | Осмотична/фізична модифікація | Краща сумісність розчину | Нестабільність механізму дії | Проникнення | Відсутність доведеного механізму | «Біоміметичне живлення» |
| Етаноламінні комплекси | Буферування pH, слабкий ліганд | Гнучкість формуляції | Сильно залежать від середовища | Нестабільність у різних умовах | pH і жорсткість води | «Нова генерація хелатів» |
| Нано-/nano-complexes | Зміна розміру часток | Висока маркетингова привабливість | Невизначеність механізмів | Кутикула та транспорт | Відсутність специфічних шляхів проникнення | «Революційне проникнення» |
Жодна форма не оптимізує одночасно всі етапи листкового живлення: стабілізацію → доставку → проникнення → транспорт → метаболічне включення. Покращення одного параметра майже завжди створює обмеження на іншому етапі. Будь-яка технологія — це компроміс між фізико-хімічними обмеженнями системи.
КОНФЛІКТ: ІНДУСТРІЯ НАЗИВАЄ ОДНЕ Й ТЕ САМЕ РІЗНИМИ СЛОВАМИ.
Сьогодні ринок мікроелементів у агробізнесі дедалі більше нагадує не систему прикладної агрохімії, а систему маркетингового ребрендингу одних і тих самих хімічних принципів. Мікроелементи подаються як високотехнологічний продукт нового покоління, де ключову роль відіграють не стільки реальні фізико-хімічні механізми, скільки складні терміни, що створюють відчуття наукового прориву. Саме тому в рекламі та презентаціях постійно з’являються фрази “нано-частинки”, “органічні транспортери”, “біоактивні комплекси”, “фізіологічно адаптовані форми”, “інтелектуальні системи доставки”, “біоміметичні технології” або “молекулярно активовані мікроелементи”. Усе це формує у фермера враження, що сучасна агрохімія вже майже навчилася керувати клітиною рослини на молекулярному рівні і створила принципово нові механізми живлення.
Але якщо прибрати маркетингову оболонку і подивитися на реальну хімію без етикетки, картина стає значно простішою і значно менш “революційною”. У більшості випадків мова йде про ті ж самі координаційні системи, які існують десятки років: класичні хелати EDTA, DTPA або EDDHA, комплекси з органічними кислотами на основі цитратів чи глюконатів, поліоли типу манітолу або сорбітолу, етаноламінні комплекси, а іноді навіть прості солі мікроелементів із додаванням поверхнево-активних речовин, стабілізаторів або регуляторів pH. Тобто фундаментальна хімія часто не змінилася — змінився лише спосіб її подачі.
Індустрія створює враження, ніби кожна нова назва означає новий біологічний механізм, хоча на практиці це часто лише інша варіація однієї і тієї ж координаційної хімії. Наприклад, якщо мікроелемент зв’язаний з органічною молекулою — його вже можуть називати “біоактивним комплексом”, хоча з точки зору хімії це може бути звичайний слабкий координаційний комплекс без унікальних транспортних властивостей. Якщо в продукті використовують поліоли або цукрові спирти, це можуть подавати як “біоміметичну систему транспорту”, хоча їхня реальна функція часто полягає лише у зміні розчинності, в’язкості або змочувальної здатності краплі. Якщо додається ПАР або регулятор поверхневого натягу — це вже можуть називати “покращеним проникненням на клітинному рівні”, хоча фактично мова йде лише про фізику поверхні листка.
Особливо показовою є тема “нано”. У багатьох випадках цей термін використовується значно ширше, ніж це допускає строгий науковий підхід. Часто достатньо просто дрібнішої дисперсії або присутності колоїдної фази, щоб продукт уже рекламувався як “нано-добриво”, хоча це не означає існування нових механізмів транспорту через кутикулу чи мембрани. Сам термін “нано” у маркетингу починає працювати не як наукова характеристика, а як психологічний символ високої технологічності.
У результаті виникає головний конфлікт сучасної агрохімії: реальна фізико-хімічна система є значно простішою і жорсткіше обмеженою законами хімії, ніж це показує маркетинг. У рослини не з’являються нові мембранні транспортери тільки тому, що на етикетці написано “біоактивний комплекс”. Кутикула не перестає бути гідрофобним бар’єром через слово “нано”. А координаційний комплекс не стає принципово новою молекулярною технологією лише через додавання органічного ліганду.
Саме тому значна частина “інновацій” на ринку мікродобрив є не революцією у фізіології рослин, а революцією у маркетинговій мові. Індустрія часто продає не нову хімію, а нову інтерпретацію старої хімії. І чим складніше звучить назва продукту, тим важче агроному відокремити реальні фізико-хімічні властивості від рекламної конструкції, побудованої навколо звичайної координаційної системи, яка існує в агрохімії вже багато десятиліть.
ЖОРСТКА НАУКА: НІЯКОЇ “ІДЕАЛЬНОЇ ФОРМИ” НЕ ІСНУЄ.
Фундаментальна проблема сучасного маркетингу мікродобрив полягає в тому, що фізіологію рослини намагаються подати як просту технологічну систему, де достатньо “сучаснішої форми”, щоб автоматично отримати краще живлення. Але рослина не працює за принципом “новіша молекула = вища ефективність”. Засвоєння мікроелементів — це складний багатоступеневий процес, який жорстко обмежений законами фізичної хімії, дифузії, мембранного транспорту та фізіології клітини. Саме тому в реальній агрохімії не існує “ідеальної форми” мікроелемента, яка могла б одночасно бути максимально стабільною, максимально проникною, максимально мобільною та максимально доступною для метаболізму. Будь-яка форма — це лише компроміс між цими властивостями.
Уся система живлення мікроелементами фактично складається з чотирьох великих етапів: стабілізації елемента в робочому розчині, доставки до поверхні рослини, проникнення через фізіологічні бар’єри та подальшого метаболічного включення всередині клітини. І саме тут починається головний конфлікт між реальною наукою та маркетинговими обіцянками.
Перший етап — стабілізація. Іон металу в розчині є хімічно нестабільною системою. Він може випадати в осад, реагувати з карбонатами, кальцієм, магнієм або іншими компонентами бакової суміші. Саме тому використовуються хелати, органічні кислоти, поліоли чи аміни — вони утримують мікроелемент у розчині та не дають йому швидко перейти в недоступну форму. Але проблема полягає в тому, що чим сильніше комплекс утримує іон, тим складніше потім рослині цей іон отримати. Надмірна стабільність — це одночасно і перевага, і обмеження.
Другий етап — доставка до поверхні листка. Тут починає працювати зовсім інша фізика: поверхневий натяг, змочування, швидкість висихання краплі, вологість повітря, температура, структура кутикули. Саме тому в продукти додають поверхнево-активні речовини, поліоли, регулятори pH та інші компоненти, які можуть покращувати фізичні властивості розчину. Але навіть якщо крапля добре розподіляється по листку, це ще не означає, що елемент реально потрапляє всередину клітини.
Третій етап — проникнення через кутикулу та мембрани. І саме тут маркетинг найчастіше починає спрощувати або навіть перекручувати фізіологію рослини. Кутикула — це складний гідрофобний бар’єр, який не має “відкритих воріт” для будь-якої органічної або нано-форми. Мембрани працюють через специфічні транспортні системи, а не через рекламні терміни. Тому сам факт того, що мікроелемент зв’язаний із манітолом, сорбітолом чи органічною кислотою, не означає автоматичного проникнення в клітину. Більше того, збільшення стабільності комплексу може навіть ускладнювати вивільнення іону біля поверхні мембрани.
Четвертий етап — метаболічне включення. Навіть після проникнення мікроелемент ще повинен перейти у фізіологічно активну форму і бути інтегрованим у ферменти, електрон-транспортні системи або інші метаболічні процеси. І тут виникає ще один парадокс: якщо комплекс надто стабільний, він може залишатися хімічно “захищеним”, але біологічно менш доступним.
Саме тому кожна спроба оптимізувати один параметр автоматично створює проблеми на іншому етапі. Якщо зробити комплекс максимально стабільним — він гірше віддаватиме іон. Якщо зробити його більш реактивним — він стане менш стабільним у баковій суміші або жорсткій воді. Якщо агресивно покращувати проникнення через кутикулу — можна втратити контроль над доставкою або підвищити ризик фітотоксичності. Це не недолік конкретної технології — це фундаментальне фізико-хімічне обмеження системи.
Але саме цей конфлікт майже ніколи не показується в маркетингу. Натомість індустрія створює ілюзію, що нова форма одночасно вирішує всі проблеми: і стабілізує, і транспортує, і проникає, і повністю засвоюється. Насправді ж жодна форма не може ідеально оптимізувати всі етапи одночасно, тому що система обмежена законами координаційної хімії, дифузії та клітинного транспорту. У результаті більшість “революційних технологій” — це не подолання обмежень, а лише інший баланс між ними.
І саме тому значна частина сучасних “інновацій” у мікроживленні є не технологічним проривом, а переформатованим продажем вічного хімічного компромісу, який існує в агрохімії вже багато десятиліть.
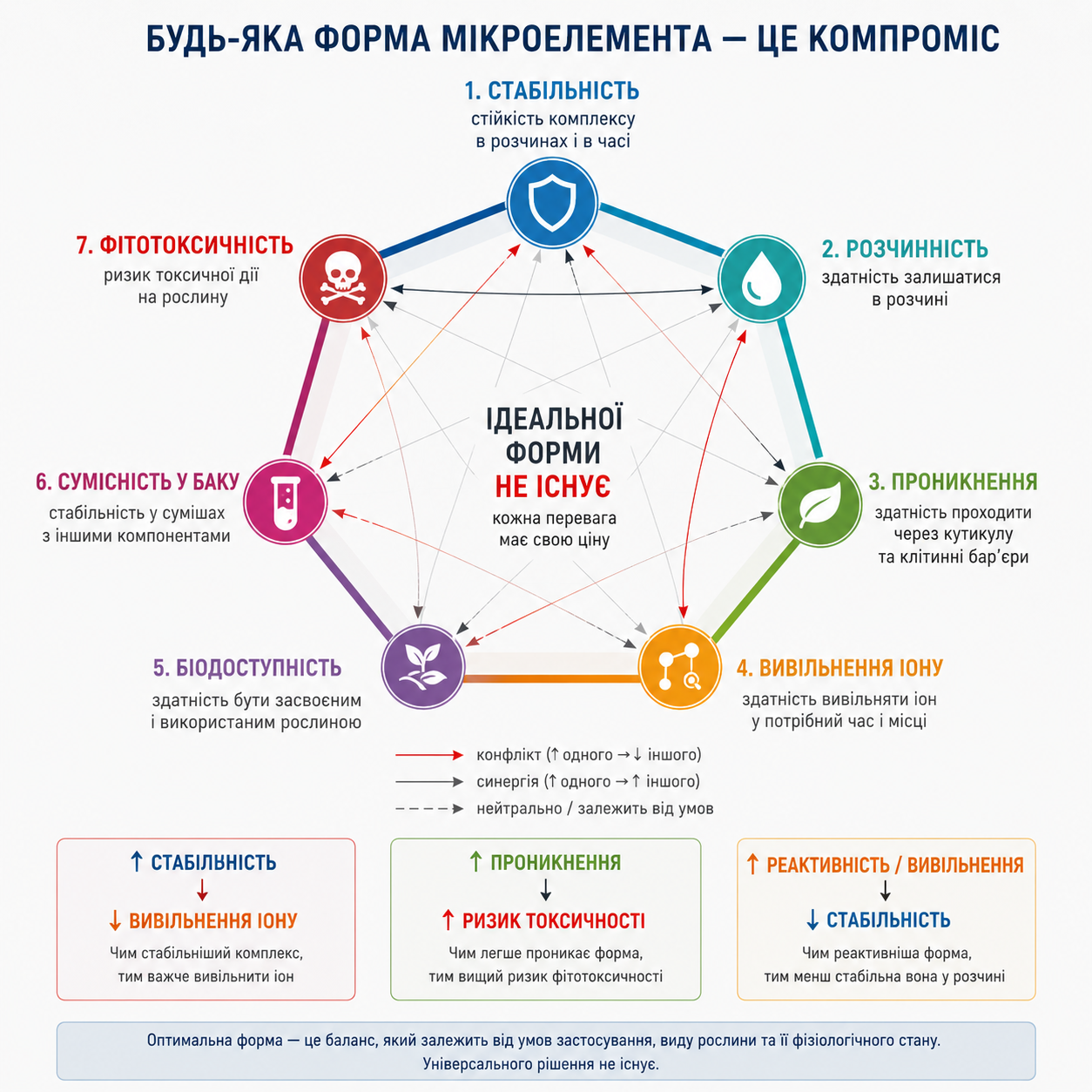 Малюнок 2. Будь-яка форма мікроелемента — це компроміс: фізико-хімічні обмеження систем мікроживлення рослин. Схематичне представлення фундаментальних компромісів між стабільністю, розчинністю, проникненням через кутикулу, швидкістю вивільнення іону, біодоступністю, сумісністю у бакових сумішах та ризиком фітотоксичності різних форм мікроелементів. Покращення одного параметра часто супроводжується погіршенням іншого через фізико-хімічні та фізіологічні обмеження системи. Рисунок демонструє, що універсальної “ідеальної форми” мікроелемента не існує, а ефективність визначається балансом властивостей у конкретних умовах застосування.
Малюнок 2. Будь-яка форма мікроелемента — це компроміс: фізико-хімічні обмеження систем мікроживлення рослин. Схематичне представлення фундаментальних компромісів між стабільністю, розчинністю, проникненням через кутикулу, швидкістю вивільнення іону, біодоступністю, сумісністю у бакових сумішах та ризиком фітотоксичності різних форм мікроелементів. Покращення одного параметра часто супроводжується погіршенням іншого через фізико-хімічні та фізіологічні обмеження системи. Рисунок демонструє, що універсальної “ідеальної форми” мікроелемента не існує, а ефективність визначається балансом властивостей у конкретних умовах застосування.
КОНФЛІКТ ХІМІЇ: КОЖНА ФОРМА — ЦЕ КОМПРОМІС.
Одна з найважливіших, але майже ніколи не озвучуваних проблем сучасної агрохімії полягає в тому, що кожна форма мікроелемента є не “ідеальним рішенням”, а лише компромісом між різними фізико-хімічними властивостями. У маркетингових матеріалах різні форми подаються як технологічні прориви або “покращені системи доставки”, але в реальності кожна з них вирішує лише окрему частину задачі і одночасно створює нові обмеження. Саме тому конфлікт між стабільністю, проникненням, вивільненням і транспортуванням є фундаментальним для всієї хімії мікроживлення.
Класичні хелати, такі як EDTA або DTPA, є хорошим прикладом цього компромісу. Їх головна перевага — висока стабільність. Вони ефективно утримують іон металу в розчині, захищають його від випадання в осад і зменшують взаємодію з кальцієм, магнієм або карбонатами у воді. Саме тому такі форми добре працюють у бакових сумішах і дозволяють зберігати мікроелемент у доступному стані навіть у складних умовах. Але ця стабільність одночасно є і проблемою. Рослині потрібен не просто стабільний комплекс, а можливість отримати сам іон у біологічно активній формі. Якщо хелат утримує метал занадто сильно, швидкість вивільнення може знижуватися, а отже частина елемента стає менш доступною для фізіологічного включення в клітинний метаболізм. Тобто перевага на етапі стабілізації перетворюється на обмеження на етапі засвоєння.
Комплекси з органічними кислотами працюють за протилежним принципом. Глюконати, цитрати або інші органічні кислоти значно слабше утримують іон металу, тому він легше вивільняється біля поверхні клітини або в апопласті. Це може прискорювати фізіологічну реакцію рослини, але одночасно робить систему менш стабільною. У жорсткій воді або складних бакових сумішах такі форми значно швидше втрачають стабільність, реагують із солями кальцію чи магнію і можуть переходити у менш доступні форми. Тобто швидкість вивільнення досягається ціною меншого контролю над стабільністю.
Поліоли та цукрові спирти — манітол, сорбітол та подібні сполуки — створюють ще один тип компромісу. У маркетингу їх часто називають “біологічними транспортерами” або “природними системами доставки”, ніби вони здатні активно переносити мікроелемент у клітину. Але в реальності їхня функція значно менш революційна. Вони переважно змінюють фізичні властивості розчину: покращують змочування поверхні листка, впливають на стабільність краплі, знижують швидкість кристалізації або покращують сумісність компонентів у баковій суміші. Це важливі технологічні функції, але вони не означають автоматичного транспорту іону через мембрани. Рослина не має універсального механізму “всмоктування поліольних комплексів”, тому більшість таких систем усе одно потребує дисоціації і переходу металу в іонну форму біля поверхні клітини.
Етаноламінні комплекси займають проміжне положення між простими солями та класичними хелатами. Етаноламін може одночасно працювати як слабкий ліганд і як регулятор pH, що покращує розчинність деяких мікроелементів. Але саме через свою проміжну природу такі системи дуже сильно залежать від умов середовища: жорсткості води, температури, кислотності та концентрації інших іонів. В одних умовах вони можуть бути достатньо стабільними, а в інших — швидко втрачати свої властивості. Тобто вони не є “ідеальною альтернативою” хелатам, а лише іншим способом балансування між стабільністю та реактивністю.
І саме тут виникає головна фізико-хімічна правда, яку маркетинг майже ніколи не озвучує: чим краще оптимізований один етап системи, тим слабшим може ставати інший. Якщо максимально посилити стабільність — погіршується вивільнення. Якщо зробити систему більш реактивною — знижується її стійкість у баковій суміші. Якщо покращити проникнення через кутикулу — можна втратити контроль над доставкою або підвищити ризик фітотоксичності. Це не проблема окремого продукту — це фундаментальне обмеження самої координаційної хімії та фізіології рослини.
Саме тому в реальній агрохімії не існує “найкращої” форми мікроелемента. Існують лише різні баланси між стабільністю, мобільністю, реактивністю і біодоступністю. Але замість пояснення цих компромісів індустрія часто створює просту ілюзію технологічного прогресу, де нова назва автоматично означає новий рівень ефективності. У результаті фермеру продають не універсальне рішення, а іншу конфігурацію старого хімічного компромісу, який у різних умовах може працювати як краще, так і гірше.
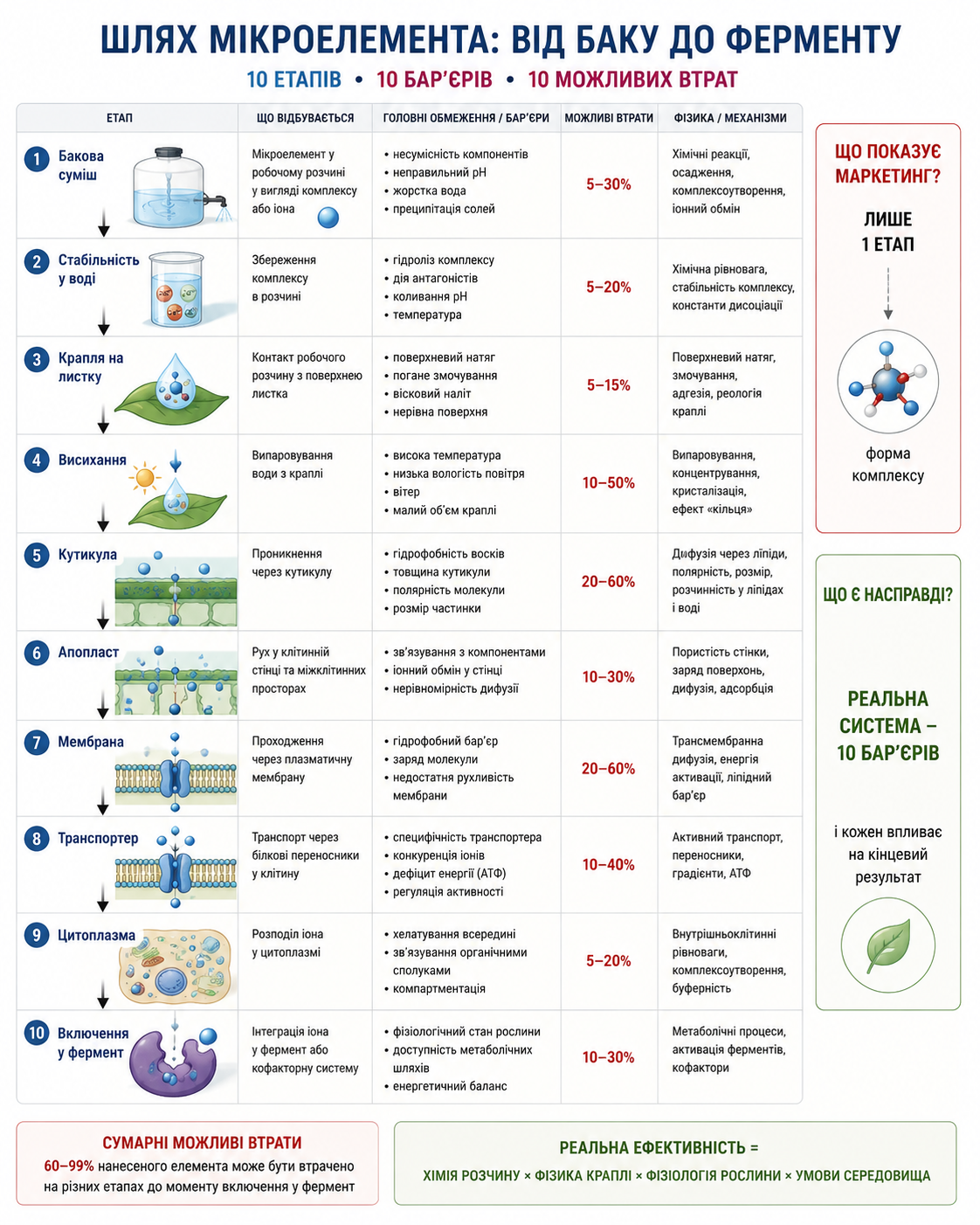
Мал. 3. Шлях мікроелемента від бакової суміші до ферментативного включення: багатоступенева система фізико-хімічних і фізіологічних бар’єрів. Схематичне представлення послідовних етапів руху мікроелемента від робочого розчину до включення у ферментативні системи рослини. Показано ключові стадії процесу: стабілізацію у баковій суміші, поведінку краплі на листковій поверхні, висихання, проходження через кутикулу, апопласт і плазматичну мембрану, транспорт через специфічні білкові переносники, внутрішньоклітинний розподіл та інтеграцію у метаболічні процеси. Для кожного етапу наведено основні фізико-хімічні обмеження, механізми втрат та фактори, що визначають ефективність засвоєння. Рисунок демонструє, що ефективність мікроживлення визначається не лише формою комплексу, а всією системою взаємодії “розчин — листок — клітина — метаболізм”.
РОЗВІНЧАННЯ МІФУ ПРО “ПРОНИКНЕННЯ В КЛІТИНУ.
Один із найпоширеніших і водночас найманіпулятивніших міфів сучасного агрохімічного маркетингу — це твердження, що “нано-форма”, “органічний комплекс” або “біоактивна молекула” автоматично забезпечують проникнення мікроелемента в клітину рослини. Саме навколо цієї ідеї побудована значна частина реклами сучасних мікродобрив і біостимуляторів. Споживачу фактично нав’язується образ рослини як простої губки, яка здатна легко поглинати будь-яку “сучасну” форму елемента, якщо вона достатньо маленька, органічна або технологічно модифікована. Але реальна фізіологія листка працює зовсім інакше.
Листкова поверхня — це не відкрита система для пасивного проникнення будь-яких молекул. Рослина еволюційно створила потужний захисний бар’єр проти неконтрольованого проникнення речовин із зовнішнього середовища. Першим таким бар’єром є кутикула — складна багатошарова структура, що складається з восків, кутину та полімерних гідрофобних компонентів. Її головна функція — не “пропускати добрива”, а навпаки, мінімізувати втрату води та захищати тканини від зовнішніх факторів. Саме тому проникнення через кутикулу визначається не маркетинговою назвою продукту, а жорсткими фізико-хімічними параметрами: розміром молекули, її зарядом, полярністю, розчинністю у водній та ліпідній фазі, а також здатністю до дифузії.
Особливо активно маркетинг спекулює на терміні “нано”. Сам факт того, що частинка має малий розмір, подається як доказ її здатності проникати в клітину. Але розмір частинки сам по собі нічого не гарантує. Навіть дуже дрібна частинка повинна пройти через гідрофобний бар’єр кутикули, клітинну стінку і плазматичну мембрану, кожна з яких має власні механізми селективності. У рослин немає універсальних “наноканалів”, через які будь-яка нано-структура автоматично потрапляє в цитоплазму. Більше того, багато наночастинок можуть агломеруватися, змінювати свій заряд або взаємодіяти з компонентами поверхні листка ще до проникнення.
Ще один поширений маркетинговий прийом — це використання поняття “органічна форма”. У рекламі часто створюється враження, що якщо мікроелемент зв’язаний із органічною молекулою — амінокислотою, поліолом, цукром чи органічною кислотою — то рослина нібито автоматично сприймає його як “природний” і легко транспортує всередину клітини. Але мембрани рослинних клітин не працюють за принципом “органічне = доступне”. Транспорт через мембрани здійснюється за допомогою специфічних білкових транспортерів, які розпізнають конкретні іони або невеликі молекули. Існування органічного комплексу ще не означає наявності механізму його прямого транспорту.
У багатьох випадках рослина взагалі не поглинає комплекс як єдину структуру. Спочатку має відбутися дисоціація — тобто вивільнення іону металу біля поверхні клітини. Саме іонна форма часто є реальною фізіологічно активною одиницею, яку розпізнають мембранні транспортери. Тому навіть якщо комплекс стабільний у баковій суміші, це не означає, що він буде ефективно включений у клітинний метаболізм.
Окремою проблемою є те, що маркетинг часто змішує поняття “контакт із листком”, “утримання на поверхні”, “проникнення в тканину” і “реальне засвоєння”. Якщо розчин краще змочує листок або довше залишається у рідкому стані, це ще не означає автоматичного транспорту в клітину. Дуже часто ефект покращення фізичних властивостей краплі подається як доказ “революційного проникнення”, хоча між цими процесами існує принципова різниця.
У результаті виникає головне спрощення сучасного агрохімічного маркетингу: складна багаторівнева система бар’єрів і селективного транспорту замінюється простою ідеєю, що достатньо зробити частинку меншою або зв’язати елемент з органічною молекулою — і проблема засвоєння буде вирішена. Але реальна фізіологія рослини не працює за такими законами. Розмір частинки не гарантує проникнення. Хімічна форма не гарантує транспорту. А “органічність” не гарантує засвоєння. Усі ці параметри лише впливають на окремі етапи складної системи, яка залишається жорстко обмеженою законами дифузії, мембранної селективності та координаційної хімії.
Мал. 4. Маркетингові концепції проти реальних фізіологічних факторів засвоєння мікроелементів рослиною. Схематичне порівняння між маркетинговими термінами, які використовуються для просування сучасних мікродобрив (“nano”, “bio”, “smart chelate”, “molecular delivery”, “physiological activation”), та реальними фізико-хімічними і фізіологічними факторами, що визначають ефективність позакореневого живлення. На відміну від маркетингових концепцій, рослина реагує не на назву технології, а на параметри середовища і власний фізіологічний стан: pH робочого розчину, вологість повітря, швидкість висихання краплі, проникність кутикули, активність мембранних транспортерів, транспірацію та рівень стресу. Рисунок ілюструє фундаментальний розрив між маркетинговою інтерпретацією “інноваційних форм” і реальною багатофакторною системою поглинання мікроелементів.
РОСЛИНА ПОГЛИНАЄ НЕ “КОМПЛЕКС”, А КОНКРЕТНИЙ ІОН АБО ОБМЕЖЕНИЙ НАБІР ФОРМ.
Ключовою фізіологічною особливістю мінерального живлення рослин є те, що більшість мембранних транспортерів еволюційно адаптовані до перенесення не “складних біоактивних комплексів”, а конкретних іонних форм елементів. Саме тому між хімічною формою добрива у препараті та реальною формою поглинання елемента рослиною існує принципова різниця. У багатьох випадках рослина поглинає не сам хелат, комплекс або “наноструктуру” як цілісну молекулярну систему, а металевий іон після часткової або повної дисоціації комплексу в ризосфері, апопласті або безпосередньо біля поверхні плазматичної мембрани. Комплекс у такому випадку виконує переважно транспортно-стабілізаційну функцію: захищає іон від осадження, окиснення, гідролізу або антагоністичних реакцій у розчині, але не обов’язково є реальною фізіологічною формою поглинання.
Це особливо важливо для розуміння сучасного ринку “нанодобрив”, “біоактивних комплексів”, “органо-мінеральних кластерів” та інших маркетингових формулювань, де складна хімічна структура часто подається як самостійна біологічно активна сутність. Насправді ж у більшості випадків кінцевою метою залишається доставка того самого іона металу до транспортних систем клітини. Рослина не має окремої еволюційно сформованої системи транспортерів для “нанохелатів”, “металоорганічних супрамолекулярних структур” або “біоактивних кластерів” як універсального класу речовин. Натомість основні транспортні білки розпізнають конкретні заряджені форми елементів, які існують у ґрунтовому розчині або апопласті.
Для заліза основними фізіологічно релевантними формами є Fe²⁺ або, у випадку стратегії II у злаків, комплекси Fe³⁺-фітосидерофорів, які мають спеціалізовані транспортери. Для цинку транспортери родини ZIP переважно працюють із Zn²⁺. Для марганцю критичною формою є Mn²⁺. Для міді — Cu²⁺ або Cu⁺ після редукції на поверхні мембрани. Бор поглинається переважно у вигляді борної кислоти або боратних форм, а молібден — у вигляді молібдат-іона MoO₄²⁻. Тобто фізіологія мінерального живлення базується не на абстрактних “інноваційних комплексах”, а на дуже конкретних хімічних формах, до яких адаптовані мембранні переносники, редуктази, канали та системи внутрішньоклітинного транспорту.
Саме тому навіть найскладніші хелатні або наноструктуровані препарати в багатьох випадках повинні зрештою перейти у класичну іонну форму перед включенням елемента у метаболізм. Після потрапляння у ризосферу або на поверхню листка комплекс часто проходить серію процесів: часткову дисоціацію, обмін лігандами, протонування, окисно-відновні реакції, взаємодію з органічними кислотами, ексудатами кореня, полісахаридами клітинної стінки та апопластичними компонентами. У результаті реальна форма, яка контактує з транспортером, може суттєво відрізнятися від початкової “маркетингової” форми препарату.
Це не означає, що хелати або комплексні форми не мають значення. Навпаки, вони можуть суттєво впливати на стабільність, мобільність, розчинність, проникнення через кутикулу, швидкість вивільнення та сумісність елементів у робочому розчині. Однак їхня роль часто є допоміжною щодо головного фізіологічного процесу — доставки доступного іона до системи клітинного транспорту. Тому наукова оцінка ефективності таких препаратів повинна базуватися не лише на складності або “інноваційності” комплексу, а насамперед на тому, наскільки ефективно він забезпечує появу доступної для рослини іонної форми у конкретних умовах середовища.
Особливо проблематичними є маркетингові твердження про “пряме клітинне поглинання нанокомплексів”, “унікальні біоактивні металокластери” або “революційні супрамолекулярні системи живлення” без демонстрації реальних механізмів транспорту через мембрану. Для підтвердження таких заяв необхідні прямі фізіологічні докази: ідентифікація транспортерів, трасування форми елемента під час поглинання, спектроскопічне підтвердження збереження комплексу в апопласті та цитоплазмі, а також демонстрація того, що рослина дійсно поглинає саме комплекс, а не іон після його розпаду. У більшості випадків таких доказів немає, а ефект пояснюється класичними механізмами мінерального живлення, відомими ще задовго до появи термінів “нано-добриво” чи “біоактивний комплекс”.
Таким чином, принципова фізіологічна реальність полягає в тому, що рослина у більшості випадків працює не з маркетинговою назвою препарату, а з конкретною хімічною формою елемента, доступною для мембранного транспорту. Саме тому складність або “нанорозмірність” комплексу сама по собі ще не є доказом вищої біологічної ефективності. Головним критерієм залишається здатність препарату забезпечити стабільну, доступну та фізіологічно сумісну форму елемента у зоні поглинання.
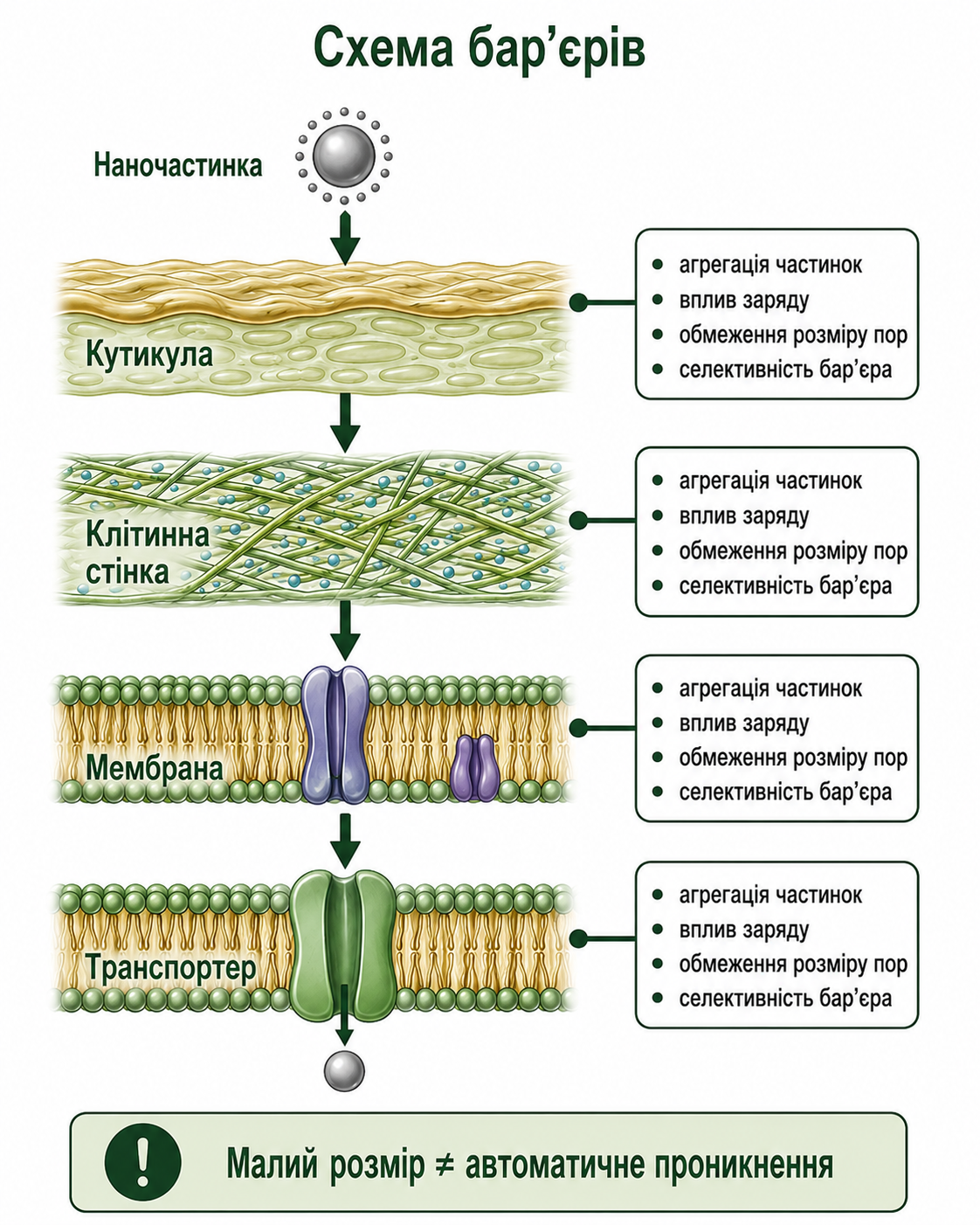
Мал. 5. Схема фізико-біологічних бар’єрів проникнення наночастинок у клітину рослини. Рисунок демонструє основні бар’єри, які обмежують проникнення наночастинок у рослинну клітину. На шляху від поверхні листка до внутрішньоклітинного транспорту наночастинки проходять через кутикулу, клітинну стінку, плазматичну мембрану та селективні транспортери. На кожному етапі виникають обмеження, пов’язані з агрегацією частинок, впливом електричного заряду, розміром пор та селективністю мембранних систем. Схема підкреслює, що малий розмір наночастинок не гарантує їх автоматичного проникнення у клітину рослини.
ПРО «ЛАБОРАТОРНУ ІЛЮЗІЮ».
Однією з найменш обговорюваних, але критично важливих проблем сучасного ринку мікродобрив, хелатів і “нанотехнологічних” препаратів є так звана “лабораторна ілюзія” ефективності. Значна частина “революційних” ефектів нових форм мікроелементів демонструється в умовах, які мають дуже обмежений зв’язок із реальним польовим середовищем. Найчастіше це гідропонні системи, стерильні субстрати, камери росту з контрольованою температурою та вологістю, очищена або деіонізована вода, стабільне освітлення, відсутність пилу, опадів, ультрафіолетового стресу та мікробного навантаження. У таких умовах навіть незначна різниця між формами мікроелемента може виглядати як “проривна технологія”. Проте саме польове середовище визначає реальну поведінку препарату, і в багатьох випадках ця поведінка суттєво відрізняється від лабораторної картини.
У контрольованих експериментах дослідник фактично ізолює препарат від більшості факторів, які в реальному полі визначають його стабільність, проникнення, утримання на листку та біологічну доступність. У лабораторії крапля робочого розчину може залишатися стабільною протягом тривалого часу, тоді як у полі вона часто висихає за лічені хвилини. У камері росту практично відсутній ультрафіолетовий спектр природного сонячного випромінювання, який здатний руйнувати органічні ліганди, фотодеградувати комплекси та змінювати хімічний стан металів. Відсутні різкі коливання температури, які впливають на швидкість гідролізу, окисно-відновні реакції та проникність кутикули. Немає пилу, воскових забруднень, сольового навантаження чи механічного пошкодження листкової поверхні. Не враховується також жорсткість польової води, де кальцій, магній і бікарбонати можуть швидко змінювати стабільність хелатів і викликати випадіння частини компонентів у менш доступні форми.
Особливо важливим фактором є мікробіом поверхні листка та ризосфери, який у лабораторних системах часто або повністю відсутній, або штучно спрощений. У реальному полі поверхня рослини є складною екосистемою, заселеною бактеріями, дріжджами, грибами та іншими мікроорганізмами, які активно взаємодіють із нанесеними речовинами. Частина органічних лігандів може швидко деградуватися мікроорганізмами ще до проникнення елемента у тканини. Деякі комплекси можуть ставати субстратом для мікробного метаболізму, а не для живлення рослини. Крім того, ексудати мікробіому можуть змінювати pH мікросередовища, редокс-стан металів і стабільність комплексів безпосередньо на поверхні листка.
Ще одним джерелом “лабораторної ілюзії” є використання модельних систем із надмірно спрощеною фізикою нанесення. У реальному полі поведінка краплі визначається десятками факторів: швидкістю вітру, температурою листка, вологістю повітря, товщиною кутикулярного воску, кутом контакту краплі, випаровуванням, кристалізацією солей після висихання та повторним зволоженням росою. Навіть якщо препарат демонструє покращене проникнення в умовах лабораторної інкубації, це ще не означає, що така перевага збережеться після швидкого польового висихання або фотодеструкції ліганда. У багатьох випадках “унікальні” властивості препарату існують лише протягом короткого проміжку часу між приготуванням розчину та деградацією в польовому середовищі.
Проблема посилюється тим, що багато маркетингових матеріалів базуються саме на лабораторних або напівконтрольованих дослідженнях, де різниця між препаратами статистично збільшується через мінімізацію шуму середовища. У реальному полі домінують зовсім інші фактори: дефіцит вологи, температура, ґрунтові обмеження, стан кореневої системи, стрес гербіцидів, фітопатогени, ущільнення ґрунту, нестача органічної речовини або дисбаланс макроелементів. На фоні цих факторів різниця між “революційним нанохелатом” і класичним хелатом або навіть неорганічною сіллю часто різко зменшується або стає статистично нестабільною.
Саме тому багато технологій, які в лабораторії демонструють суттєве підвищення поглинання, біодоступності або швидкості проникнення, у польових умовах втрачають більшу частину своєї “унікальності”. Це не обов’язково означає, що препарат не працює взагалі, але означає, що реальна агрономічна цінність може бути значно меншою за маркетингові обіцянки. Ефект, який у стерильній системі виглядає як “зростання ефективності на 40–60 %”, у полі може перетворюватися на кілька відсотків різниці, які перебувають у межах природної варіабельності врожайності.
Тому для об’єктивної оцінки сучасних форм мікроелементів критично важливими є не лише лабораторні показники, а насамперед багаторічні польові дослідження в реальних агрономічних умовах. Лише польове середовище здатне показати, наскільки технологія є стабільною під дією ультрафіолету, коливань температури, деградації краплі, мікробної активності, жорсткої води та інших факторів, які фактично визначають долю препарату після внесення. Без такого тестування багато “інноваційних” ефектів залишаються переважно артефактом контрольованої лабораторної системи, а не доказом принципово нової ефективності в реальному землеробстві.
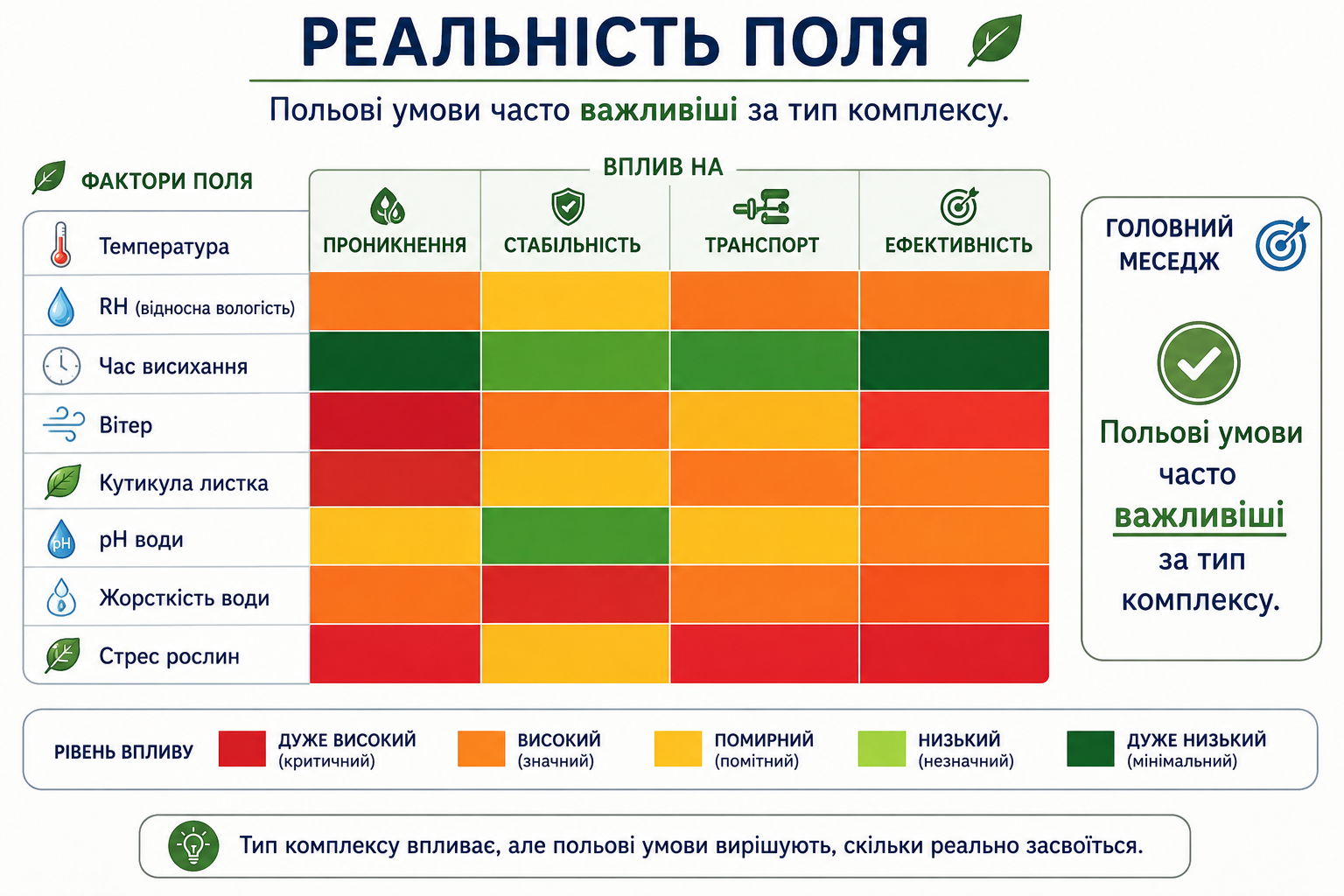
Мал. 6. Реальність поля: вплив польових умов на ефективність засвоєння. Heatmap-схема відносного впливу польових факторів (температура, відносна вологість, час висихання, вітер, стан кутикули, pH і жорсткість води, фізіологічний стрес рослин) на проникнення, стабільність, транспорт і загальну ефективність засвоєння речовин. Показано, що реальні польові умови часто мають більший вплив на результат, ніж тип координаційного комплексу.
БІОСТИМУЛЯЦІЯ НЕ Є ЖИВЛЕННЯМ.
Однією з найпоширеніших причин неправильного трактування ефективності сучасних мікродобрив є змішування двох принципово різних явищ — мінерального живлення та біостимуляції. У багатьох випадках позитивна реакція рослини після внесення препарату автоматично пояснюється “кращим засвоєнням мікроелемента”, хоча реальний механізм ефекту може бути зовсім іншим. Це особливо характерно для продуктів, які, крім мікроелементів, містять амінокислоти, поліоли, органічні екстракти, поверхнево-активні речовини, антистресові компоненти, осмопротектори або різні сигнальні молекули. Такі речовини здатні швидко змінювати фізіологічний стан рослини навіть без суттєвого збільшення реального надходження мінерального елемента у метаболізм.
Біостимуляція і живлення — це не тотожні процеси. Мінеральне живлення передбачає забезпечення рослини необхідними елементами у кількостях, достатніх для структурних і ферментативних функцій. У цьому випадку ключовим є надходження конкретного елемента — цинку, марганцю, бору, міді, молібдену чи заліза — у фізіологічно доступній формі до клітинних транспортних систем і подальше включення в метаболічні процеси. Натомість біостимуляція часто працює через короткочасну зміну фізіологічного стану рослини: регуляцію осмотичного балансу, антиоксидантний захист, сигнальні каскади, зміну проникності мембран, стимуляцію фотосинтетичної активності або пом’якшення стресових реакцій. У таких випадках візуальний ефект може бути дуже помітним навіть за мінімального внеску самого мікроелемента.
Саме тому препарати з амінокислотами, поліолами або екстрактами часто викликають швидке “позеленіння” рослини, покращення тургору, тимчасову активізацію росту чи візуальне зменшення симптомів стресу. Амінокислоти можуть виступати джерелом легкодоступного органічного азоту або сигнальними молекулами для метаболічних процесів. Поліоли здатні працювати як осмопротектори, стабілізуючи клітинні структури під час посухи або високої температури. Морські екстракти часто містять фітогормоноподібні сполуки, антиоксиданти та органічні метаболіти, що впливають на фізіологію рослини незалежно від мікроелементного живлення. Поверхнево-активні речовини можуть тимчасово покращувати змочування листка та проникнення компонентів через кутикулу. Антистресові добавки можуть знижувати інтенсивність оксидативного стресу або підтримувати роботу мембран у несприятливих умовах.
У результаті рослина дійсно може виглядати “краще” після внесення такого препарату, але це ще не є автоматичним доказом того, що саме мікроелемент засвоївся ефективніше або що його фізіологічна доступність принципово зросла. Дуже часто короткостроковий позитивний ефект пов’язаний саме з біостимулюючими компонентами, а не з унікальною формою цинку, марганцю чи бору. Це особливо важливо, тому що візуальна реакція рослини є психологічно дуже переконливою для агронома або фермера. Якщо через кілька днів після обробки листок стає темнішим, рослина виглядає “живішою”, а тургор покращується, це легко інтерпретується як доказ “надзвичайного засвоєння” мікроелемента, навіть якщо реальний механізм полягає у тимчасовій фізіологічній стимуляції або антистресовій дії.
Проблема посилюється тим, що маркетинг часто навмисно об’єднує ці різні механізми в одну концепцію. Біостимулюючий ефект препарату подається як доказ “революційної системи доставки мікроелементів”, хоча в реальності рослина може реагувати насамперед на органічні допоміжні компоненти. У деяких випадках позитивний ефект після внесення взагалі може бути пов’язаний переважно з ПАР, які покращують змочування листка, або з антистресовими сполуками, що тимчасово знижують наслідки посухи чи температурного стресу. Це не означає, що препарат є “поганим” або “непрацюючим”, але означає, що механізм його дії необхідно інтерпретувати коректно.
Особливо важливо розділяти ці поняття при оцінці економічної доцільності продукту. Якщо препарат дає короткочасний косметичний або антистресовий ефект, це ще не означає, що він вирішує системний дефіцит елемента живлення. Рослина може тимчасово покращити зовнішній вигляд навіть за збереження фундаментальної нестачі цинку, бору чи марганцю в метаболічно важливих тканинах. Візуальне “оздоровлення” не завжди еквівалентне реальному усуненню дефіциту або стабільному підвищенню продуктивності.
Саме тому в сучасній агрохімії критично важливо відокремлювати ефекти біостимуляції від ефектів мінерального живлення. Якщо продукт містить мікроелементи разом із амінокислотами, екстрактами чи антистресантами, необхідно оцінювати, яка частина ефекту пов’язана з реальним надходженням елемента, а яка — з короткостроковою фізіологічною стимуляцією. Без такого розділення будь-яке “позеленіння” або покращення тургору легко перетворюється на маркетинговий аргумент про “надзвичайну біодоступність”, хоча реальна фізіологія процесу може бути значно складнішою та менш сенсаційною.
ДОЗА, КОНЦЕНТРАЦІЯ ТА ЕФЕКТ НАСИЧЕННЯ: ЧОМУ “СУПЕРФОРМА” НЕ СКАСОВУЄ ФІЗІОЛОГІЮ.
Однією з найбільш недооцінених тем у сучасному маркетингу мікродобрив є фундаментальна роль дози, концентрації та ефекту насичення. У багатьох рекламних концепціях основна увага зосереджується майже виключно на “унікальній формі” елемента: нанокомплексі, біохелаті, супрамолекулярній структурі чи “інтелектуальній системі доставки”. Проте в реальній фізіології рослини сама форма є лише одним із факторів ефективності. Не менш важливими є кількість елемента, його локальна концентрація, швидкість доставки, тривалість доступності та співвідношення між дефіцитом і токсичністю. Навіть найстабільніший і найдорожчий комплекс не працюватиме, якщо рослина просто не отримає достатньої кількості необхідного елемента. І навпаки — надмірна концентрація може викликати токсичність, пошкодження тканин або метаболічний дисбаланс незалежно від “м’якості” чи складності ліганду.
Мінеральне живлення рослин підпорядковується базовим законам біохімії та транспорту речовин. Для кожного елемента існує певний фізіологічний діапазон концентрацій: нижче нього виникає дефіцит, у межах оптимуму забезпечується нормальна робота ферментів і метаболічних систем, а вище — зростає ризик токсичності. Цей принцип не зникає лише тому, що елемент перевели у “наноформу” або зв’язали з органічним лігандом. Якщо доза цинку, бору чи міді є нижчою за фізіологічний поріг потреби культури, жодна “революційна система доставки” не здатна повністю компенсувати елементарний дефіцит кількості.
Особливо важливо розуміти, що багато маркетингових тверджень фактично намагаються створити враження, ніби “суперефективна форма” дозволяє майже скасувати поняття дози. Часто можна зустріти заяви про “ультранизькі норми внесення”, “надзвичайну біодоступність” або “максимальну ефективність кожної молекули”. Частково це може бути правдою в умовах, коли препарат справді покращує проникнення або зменшує втрати елемента. Однак навіть за ідеального поглинання рослина все одно потребує певної абсолютної кількості атомів елемента для побудови ферментів, білків, мембранних систем і метаболічних структур. Неможливо повністю компенсувати нестачу кількості лише складністю хімічної форми.
Не менш важливим є явище saturation effect — ефекту насичення транспортних систем. Більшість мембранних транспортерів мають обмежену швидкість роботи. Коли концентрація елемента досягає певного рівня, подальше збільшення кількості речовини вже не призводить до пропорційного збільшення поглинання. Це фундаментальний принцип ферментативної кінетики та мембранного транспорту. У результаті навіть якщо препарат забезпечує дуже високу локальну концентрацію мікроелемента на поверхні листка або кореня, рослина не здатна безмежно збільшувати швидкість його засвоєння. Частина елемента може залишатися невикористаною, осаджуватися, вимиватися або переходити у форми, що вже не беруть участі у живленні.
Паралельно з цим надмірна концентрація мікроелементів може створювати локальну токсичність. Це особливо характерно для позакореневих обробок, де висока концентрація солей або металів у краплі здатна викликати дегідратацію клітин, пошкодження кутикули, окислювальний стрес або некротичні опіки тканин. Навіть “м’які” хелатні форми не гарантують повної безпечності при перевищенні концентрації. Металевий іон залишається біологічно активним, а при надлишку може порушувати роботу ферментів, створювати дисбаланс інших елементів або генерувати реактивні форми кисню. Особливо вузьке “вікно” між дефіцитом і токсичністю мають мідь, бор та марганець.
Крім абсолютної дози, велике значення має швидкість доставки елемента. Якщо надходження відбувається занадто швидко, рослина може не встигати інтегрувати елемент у метаболізм, і локальна концентрація стає токсичною. Якщо ж вивільнення надто повільне, то навіть стабільний комплекс може не покривати потреби культури у критичні фази розвитку. Таким чином, ефективність препарату визначається не лише тим, “у що загорнутий” мікроелемент, а й тим, скільки елемента реально надходить, з якою швидкістю та чи відповідає ця динаміка фізіологічним потребам рослини.
Особливо показовим є те, що в польових умовах фактор дози часто має більший вплив, ніж тип ліганду. У багатьох випадках адекватна кількість простого сульфату або класичного хелату може забезпечувати кращий результат, ніж дуже складний “нанобіокомплекс”, внесений у недостатній нормі. Саме тому порівняння препаратів без урахування еквівалентної кількості діючого елемента є науково некоректним. Якщо один продукт містить у кілька разів менше реально доступного цинку чи бору, його “вища технологічність” не обов’язково компенсує дефіцит абсолютної кількості елемента.
Таким чином, реальна ефективність мікродобрив визначається не лише складністю хімічної форми, а балансом між доступністю, дозою, концентрацією, швидкістю доставки та фізіологічними межами поглинання. Жодна “суперформа” не скасовує фундаментальних законів транспорту речовин і метаболізму. Рослина все одно працює в межах біохімічних порогів, де недостатня кількість не забезпечує ефекту, а надлишок переходить у токсичність. Саме тому при оцінці будь-якого “інноваційного” препарату необхідно аналізувати не лише його маркетингово описану форму, а й реальну дозу елемента, концентрацію в робочому розчині, ризики локальної токсичності та відповідність фізіологічним потребам культури.
ЕФЕКТ НЕ Є ДОКАЗОМ МЕХАНІЗМУ: ПРОБЛЕМА ВІДСУТНОСТІ ПРЯМИХ ФІЗІОЛОГІЧНИХ ДОКАЗІВ
Однією з найбільших проблем сучасного ринку “інноваційних” мікродобрив, нанокомплексів і біоактивних систем доставки є відсутність прямих фізіологічних доказів заявлених механізмів дії. Більшість компаній активно демонструють кінцеві польові або візуальні ефекти: підвищення врожайності, покращення NDVI, інтенсивніше забарвлення листка, швидший старт культури, кращий тургор або зменшення симптомів стресу. Проте такі показники самі по собі ще не є доказом того, що препарат працює саме через механізм, який декларується в маркетингових матеріалах. У багатьох випадках спостережуваний ефект і реальний механізм дії — це різні речі.
Саме тут виникає одна з ключових наукових проблем: більшість “революційних” технологій практично не супроводжуються прямими дослідженнями транспорту й долі елемента в рослині. Дуже рідко публікуються роботи з використанням ізотопного трасування, яке дозволяє точно відстежити шлях конкретного елемента від препарату до тканин рослини. Майже відсутні confocal imaging дослідження, здатні показати, чи справді “нанокомплекс” проникає через кутикулу та мембрану як цілісна структура. Рідко демонструються transporter studies — експерименти, які підтверджують взаємодію препарату з конкретними мембранними переносниками. Практично не публікуються детальні real uptake kinetics — дослідження швидкості та динаміки реального поглинання елемента у часі.
Натомість у більшості випадків компанії показують лише кінцевий агрономічний результат. Але врожайність, NDVI або зелений колір листка не можуть автоматично підтвердити заявлений механізм доставки елемента. Якщо після внесення препарату рослина стала “зеленішою”, це ще не означає, що саме “супрамолекулярний нанокомплекс” проник у клітину в незмінному вигляді. Ефект може бути пов’язаний із десятками інших факторів: біостимуляцією, антистресовою дією, ПАР-компонентами, амінокислотами, покращенням змочування листка або просто внесенням додаткової кількості елемента незалежно від його “унікальної” форми.
Це фундаментальна проблема сучасної агрохімічної комунікації: кінцевий ефект часто використовується як непрямий доказ механізму, хоча в науці такі речі не є еквівалентними. Те, що препарат працює, ще не означає, що він працює саме так, як це заявляється. У фізіології рослин причинно-наслідковий зв’язок між формою елемента, способом транспорту та кінцевим ефектом повинен підтверджуватися окремо. Без цього твердження про “унікальне клітинне проникнення”, “інтелектуальну систему доставки”, “нанотранспорт через мембрану” або “біоактивні металокластери” залишаються переважно гіпотезами або маркетинговими інтерпретаціями.
Особливо показовим є те, що для багатьох продуктів відсутні докази збереження комплексу після контакту з реальним польовим середовищем. Невідомо, чи існує заявлена “наноструктура” після змішування з жорсткою водою, після висихання краплі, під дією ультрафіолету або після контакту з поверхнею листка. Ще менш зрозуміло, чи залишається цей комплекс стабільним у апопласті, чи не розпадається до класичних іонних форм ще до моменту контакту з транспортером. Без таких даних твердження про “революційний транспортний механізм” залишаються значною мірою спекулятивними.
У сучасній молекулярній біології та фізіології рослин існує великий набір інструментів, які дозволяють реально перевіряти подібні механізми. Ізотопне мічення може показати точний шлях елемента. Флуоресцентне та конфокальне зображення здатне візуалізувати локалізацію частинок і комплексів у тканинах. ICP-MS, XANES, synchrotron spectroscopy та інші методи дозволяють визначати хімічний стан металу після поглинання. Генетичні дослідження транспортерів можуть демонструвати, чи залежить ефект препарату від конкретних білків переносу. Саме такі дані є реальним механістичним доказом у сучасній науці, а не лише статистично значуща різниця врожайності.
Відсутність прямих доказів не означає автоматично, що препарат неефективний. Продукт може реально покращувати живлення або фізіологічний стан рослини. Однак без механістичних досліджень неможливо достовірно стверджувати, що ефект обумовлений саме тим “унікальним” механізмом, який використовується у маркетинговому позиціонуванні. Це принципово важливе розмежування між агрономічним результатом і біологічним поясненням цього результату.
Саме тому критичне наукове мислення вимагає відокремлювати два рівні доказів. Перший рівень — це факт наявності ефекту: підвищення врожайності, покращення росту чи антистресова дія. Другий рівень — це доказ механізму: яким саме шляхом цей ефект виник, у якій формі елемент поглинався, чи брав участь заявлений комплекс у транспорті, і чи справді “інноваційна форма” працювала так, як це декларується. У сучасному ринку мікродобрив перший тип доказів використовується дуже активно, тоді як другий часто майже відсутній.
Саме ця різниця між “ефектом” і “доказом механізму” є одним із найсильніших наукових аргументів у критичному аналізі сучасних агрохімічних технологій. Без прямих фізіологічних і молекулярних підтверджень багато тверджень про “революційні” форми мікроелементів залишаються значною мірою маркетинговими концепціями, а не повноцінно доведеними біологічними механізмами.
ТЕРМОДИНАМІКА ПРОТИ КІНЕТИКИ: ЧОМУ СТАБІЛЬНІСТЬ КОМПЛЕКСУ ЩЕ НЕ ОЗНАЧАЄ ЕФЕКТИВНІСТЬ
Однією з найглибших і водночас найменш обговорюваних проблем сучасних мікродобрив є різниця між термодинамічною стабільністю комплексу та його реальною кінетичною поведінкою в біологічному середовищі. У більшості маркетингових описів препарати оцінюються дуже спрощено: “стабільний хелат” автоматично подається як кращий, сучасніший і ефективніший. Проте в реальній фізичній хімії та фізіології рослин стабільність — це лише частина проблеми. Не менш важливими є швидкість дисоціації комплексу, кінетика обміну лігандів, динаміка переходу металу між різними хімічними формами та час, протягом якого елемент залишається доступним для транспортних систем рослини.
Термодинамічна стабільність описує, наскільки energetically вигідним є існування певного комплексу в рівноважному стані. Високий stability constant означає, що метал міцно зв’язаний із лігандом і комплекс має низьку схильність до спонтанного розпаду. Саме на цьому часто будується маркетинг “суперстабільних” хелатів або нанокомплексів. Однак рослина працює не в абстрактній рівноважній системі, а в динамічному середовищі, де ключову роль відіграє кінетика — тобто швидкість хімічних процесів у реальному часі.
Для ефективного живлення недостатньо, щоб комплекс був просто стабільним. Метал повинен вивільнятися з комплексу з такою швидкістю, яка відповідає фізіологічним потребам рослини. Якщо дисоціація надто повільна, транспортні системи можуть просто не отримувати достатньої кількості доступного іона у критичний момент. У такому випадку термодинамічно “ідеальний” комплекс фактично поводиться як резервуар металу, який занадто повільно взаємодіє з біологічною системою. Навпаки, надто швидка дисоціація може призводити до втрати переваг комплексу: осадження металу, реакцій з антагоністами, окиснення або локальної токсичності.
Саме тому в реальних системах надзвичайно важливими стають динаміка обміну лігандів — процеси обміну лігандів між металом і компонентами середовища. Після потрапляння препарату в бакову суміш, на поверхню листка або в ризосферу комплекс починає взаємодіяти з величезною кількістю інших молекул: органічними кислотами, амінокислотами, фосфатами, карбонатами, ексудатами кореня, компонентами клітинної стінки та мікробними метаболітами. У результаті метал постійно переходить між різними координаційними станами. Комплекс, який у лабораторній пробірці виглядає “стабільним”, у реальному польовому середовищі може швидко втрачати початкову структуру через конкуренцію лігандів і зміну хімічних умов.
Особливо важливо, що рослина сама активно втручається у цю динаміку. Корені виділяють протони, органічні кислоти та редукуючі агенти, здатні змінювати координаційний стан металів. На поверхні мембрани працюють ферменти, які можуть відновлювати або окиснювати метал перед транспортом. У результаті поглинання елемента часто визначається не абсолютною стабільністю початкового комплексу, а швидкістю та характером його перебудови в мікросередовищі ризосфери або апопласту.
Це створює фундаментальний парадокс сучасних хелатних систем: надмірна стабільність не завжди є перевагою. Якщо комплекс занадто інертний, рослина може мати труднощі з вилученням металу для метаболічного використання. Саме тому деякі “слабші” комплекси іноді демонструють кращу фізіологічну ефективність, ніж дуже міцні хелати. Їхня перевага може полягати не у вищій стабільності, а у кращому балансі між стабілізацією та швидкістю вивільнення металу.
Додатково ситуацію ускладнює те, що кінетика хімічних процесів сильно залежить від реального польового середовища. Температура, pH, жорсткість води, ультрафіолет, концентрація солей, швидкість висихання краплі та активність мікробіому можуть радикально змінювати швидкість ligand exchange reactions і дисоціації комплексу. Тому поведінка препарату в лабораторній системі з контрольованим pH і стерильним середовищем часто не відображає його реальної динаміки у полі.
Саме через це спрощене протиставлення “стабільний = хороший” і “нестабільний = поганий” є науково недостатнім. У фізіології рослин критично важливий не лише термодинамічний стан комплексу, а його динамічна поведінка у часі. Реальна ефективність визначається тим, наскільки швидко й у якій формі метал переходить між різними координаційними станами, як він взаємодіє з транспортними системами та чи відповідає ця кінетика потребам рослини в конкретний момент розвитку.
Таким чином, сучасні мікродобрива слід оцінювати не лише за “силою зв’язування” металу, а як динамічні хімічні системи, де головну роль відіграє баланс між стабільністю, мобільністю, швидкістю обміну лігандів і кінетикою вивільнення елемента. Саме ця різниця між термодинамікою та кінетикою пояснює, чому одна й та сама форма металу може поводитися зовсім по-різному в лабораторії, у баковій суміші, на листковій поверхні та всередині живої рослини.
АПОПЛАСТ ПРОТИ СИМПЛАСТУ: ЧОМУ ДОСТАВКА ДО ПОВЕРХНІ КЛІТИНИ ЩЕ НЕ ОЗНАЧАЄ РЕАЛЬНЕ ПОГЛИНАННЯ
Однією з ключових, але часто ігнорованих тем у сучасному обговоренні мікродобрив є принципова різниця між рухом елемента в апопласті та його реальним входженням у симпласт через плазматичну мембрану. У багатьох маркетингових матеріалах ці процеси фактично змішуються: якщо препарат проник через кутикулу або перемістився в тканині листка, це автоматично подається як доказ “ефективного засвоєння”. Проте з точки зору фізіології рослин це лише частина шляху. Переміщення елемента в апопластичному просторі ще не означає, що він був реально поглинутий клітиною та включений у метаболізм.
Апопласт — це позаклітинний простір рослини, який включає клітинні стінки, міжклітинники та систему капілярів, заповнених водою і розчиненими речовинами. Це своєрідна “зовнішня транспортна мережа” рослини, де вода, іони та різні молекули можуть переміщуватися без перетину плазматичної мембрани. Натомість симпласт — це внутрішній безперервний простір живих клітин, з’єднаних плазмодесмами, де речовини вже перебувають під прямим контролем клітинного метаболізму. Саме перехід через плазматичну мембрану з апопласту в симпласт є справжнім актом фізіологічного поглинання.
Ця різниця має фундаментальне значення для оцінки ефективності мікродобрив. Багато препаратів можуть досить успішно переміщуватися в апопласті: проникати через кутикулу, розподілятися між клітинними стінками, накопичуватися у міжклітинниках або навіть рухатися разом із водним потоком. Але якщо елемент не проходить через мембрану до симпласту, він не стає повноцінною частиною внутрішньоклітинного метаболізму. Рослина фактично може мати значну кількість металу в апопласті, але при цьому продовжувати відчувати функціональний дефіцит усередині клітин.
Саме мембрана є головним фізіологічним бар’єром і точкою селекції. Транспорт через неї здійснюється не хаотично, а через специфічні білкові системи: транспортери, канали, переносники та редуктази. Більшість із них працює з конкретними іонними формами металів — Fe²⁺, Zn²⁺, Mn²⁺, Cu⁺/Cu²⁺, молібдатом або боратними формами. Це означає, що навіть якщо “нанокомплекс” або хелат успішно досягає апопласту, рослина все одно часто повинна або частково дисоціювати комплекс, або перебудувати його хімічно перед реальним транспортом через мембрану.
У багатьох маркетингових описах відбувається підміна понять між “проникненням” і “поглинанням”. Наприклад, якщо флуоресцентно мічена частинка або комплекс виявляється в тканині листка, це ще не означає, що він перетнув мембрану й потрапив у симпласт живих клітин. Дуже часто такі структури залишаються локалізованими саме в апопластичному просторі або адсорбуються на клітинних стінках. З фізіологічної точки зору це принципово різні стани. Апопласт може виступати тимчасовим резервуаром елементів, але не гарантує їхнього метаболічного використання.
Особливо важливо це для металів, які сильно взаємодіють із компонентами клітинної стінки. Пектини та карбоксильні групи клітинних стінок мають високу здатність зв’язувати катіони, особливо кальцій, мідь, цинк і залізо. У результаті значна частина внесеного елемента може затримуватися в апопласті як “іонно зв’язаний пул”, не доходячи до транспортерів плазматичної мембрани. Саме тому висока концентрація елемента в тканині ще не завжди означає його високу фізіологічну доступність.
Додатково ситуацію ускладнює те, що хімічні умови апопласту сильно відрізняються від цитоплазми. Тут інший pH, інший редокс-стан, висока концентрація полімерів клітинної стінки та активні процеси ligand exchange. У цьому середовищі комплекс може змінювати структуру, дисоціювати або переходити в інші форми ще до контакту з мембранним транспортером. Саме тому “стабільність” препарату в робочому розчині не гарантує його стабільності в апопласті.
У кореневій системі різниця між апопластичним і симпластичним шляхом є ще більш критичною. Вода й іони можуть відносно вільно рухатися по клітинних стінках кори кореня до ендодерми, але Casparian strip блокує подальший апопластичний рух. Для потрапляння у центральний циліндр і ксилему елемент повинен перетнути мембрану живих клітин. Це означає, що на певному етапі навіть найбільш мобільний комплекс усе одно змушений пройти через класичний етап мембранного транспорту.
Саме тому сучасні твердження про “унікальне проникнення” мікродобрив часто потребують значно глибшого фізіологічного підтвердження. Необхідно окремо демонструвати: чи препарат лише переміщується в апопласті, чи реально потрапляє у симпласт; чи зберігається комплекс після контакту з клітинною стінкою; чи бере участь у транспорті конкретний мембранний переносник; і в якій саме формі елемент входить у цитоплазму.
Це принципове розділення між апопластичним рухом і симпластичним поглинанням є одним із найважливіших фізіологічних аспектів сучасної агрохімії. Воно показує, що доставка елемента до поверхні клітини — це ще не завершення процесу живлення, а лише підготовчий етап перед найважливішою подією: контрольованим транспортом через мембрану в живу метаболічну систему рослини.
ПРОБЛЕМА ПОЛЯРНОСТІ КУТИКУЛИ: ЧОМУ НЕ ІСНУЄ “ІДЕАЛЬНОЇ” ФОРМИ ДЛЯ ПОЗАКОРЕНЕВОГО ПОГЛИНАННЯ
Однією з найскладніших і водночас найменш інтуїтивних проблем позакореневого живлення є так звана проблема полярності кутикули. Саме вона пояснює, чому створення “ідеальної” форми мікроелемента для листкового проникнення є набагато складнішим завданням, ніж це часто подається в маркетингових концепціях. Кутикула рослини не є ані повністю гідрофобною мембраною, ані простою водною плівкою. Це складна багатофазна структура, у якій одночасно співіснують ліпофільні та гідрофільні домени. Саме ця подвійна природа створює фундаментальний фізико-хімічний парадокс: надто полярні молекули мають труднощі з проходженням через гідрофобні компоненти кутикули, тоді як надто ліпофільні молекули погано рухаються через водні мікропори та полярні канали.
Кутикула складається переважно з кутину, восків і ліпідних компонентів, які формують гідрофобний бар’єр для втрати води. Саме ця властивість забезпечує рослині захист від висихання. Проте кутикула не є суцільною непроникною “пластиковою плівкою”. У її структурі існують водні мікродомени, полярні канали та нанопори, через які можуть переміщуватися вода й розчинені речовини. У результаті транспорт через кутикулу визначається не одним механізмом, а складною взаємодією між ліпофільними та гідрофільними шляхами проникнення.
Саме тут виникає фундаментальна проблема для мікродобрив. Іонні форми металів — Zn²⁺, Mn²⁺, Fe²⁺, Cu²⁺ — є високополярними й добре взаємодіють із водними порами, але мають труднощі з проходженням через ліпофільні воскові ділянки кутикули. Через це прості солі часто демонструють обмежене проникнення через листкову поверхню, особливо за швидкого висихання краплі або товстого воскового шару.
На перший погляд здається логічним “вирішити” цю проблему шляхом створення більш ліпофільних комплексів. Саме тому багато сучасних препаратів використовують органічні ліганди, амінокислоти, поліоли або інші компоненти, які підвищують ліпофільність системи. Частково це справді може покращувати взаємодію з кутикулярними восками. Однак тут виникає протилежна проблема: надто ліпофільні молекули можуть добре адсорбуватися в кутикулі, але погано рухатися через водні мікроканали та апопластичне середовище. Вони ніби “застрягають” у ліпідних фазах кутикули або повільно виходять із них.
Таким чином, система стикається з фундаментальним компромісом. Надто полярні структури погано проходять через гідрофобний бар’єр, а надто ліпофільні — мають труднощі з подальшим переміщенням у водному середовищі клітинних стінок і апопласту. Саме тому не існує універсально “ідеальної” форми мікроелемента для позакореневого проникнення. Будь-яка система є компромісом між водорозчинністю, ліпофільністю, стабільністю, швидкістю вивільнення та мобільністю в різних фазах листкової поверхні.
Ситуацію ще більше ускладнює те, що властивості кутикули постійно змінюються залежно від виду рослини, віку листка, освітлення, температури, вологості та стресових умов. Молоді листки мають одну структуру кутикулярних восків, старі — іншу. За посухи кутикула часто стає більш гідрофобною та менш проникною. Температура змінює в’язкість воскових шарів. Вологість впливає на гідратацію водних мікропор. У результаті форма, яка добре працює в одних умовах, може різко втрачати ефективність в інших.
Особливо показовим є те, що багато “суперпроникних” систем демонструють ефект лише в умовах тривалого змочування листка або високої вологості. Коли крапля швидко висихає у польових умовах, транспорт через водні канали кутикули різко обмежується. Якщо ж препарат надто ліпофільний, він може залишатися локалізованим у восковому шарі без ефективного переходу до живих тканин. Саме тому лабораторні результати проникнення часто погано відтворюються у реальному полі.
Ця проблема також пояснює, чому поверхнево-активні речовини та ад’юванти іноді мають більший вплив на ефективність позакореневого живлення, ніж сама форма мікроелемента. Вони можуть тимчасово змінювати змочування поверхні, гідратацію кутикули, розподіл краплі або проникність певних доменів. Але навіть вони не усувають фундаментального фізико-хімічного компромісу між полярністю та ліпофільністю.
У результаті сучасні мікродобрива для листкового внесення фактично працюють у дуже вузькому “вікні” фізико-хімічного балансу. Система повинна бути достатньо полярною для руху у водних мікроканалах, але водночас достатньо ліпофільною для взаємодії з кутикулярними восками. Вона має бути стабільною, але не надто інертною; рухливою, але не надто швидко вимиватися або кристалізуватися. Саме тому твердження про створення “універсальної ідеальної форми” мікроелемента суперечить самій фізичній природі кутикули.
Проблема полярності кутикули є ще одним фундаментальним доказом того, що ефективність позакореневого живлення визначається не однією “чарівною” формою, а складним балансом фізики поверхні, хімії комплексів, динаміки краплі, стану кутикули та фізіології рослини. Саме цей баланс, а не сама по собі “нанотехнологічність” чи складність комплексу, визначає реальне проникнення елемента в живу тканину рослини.
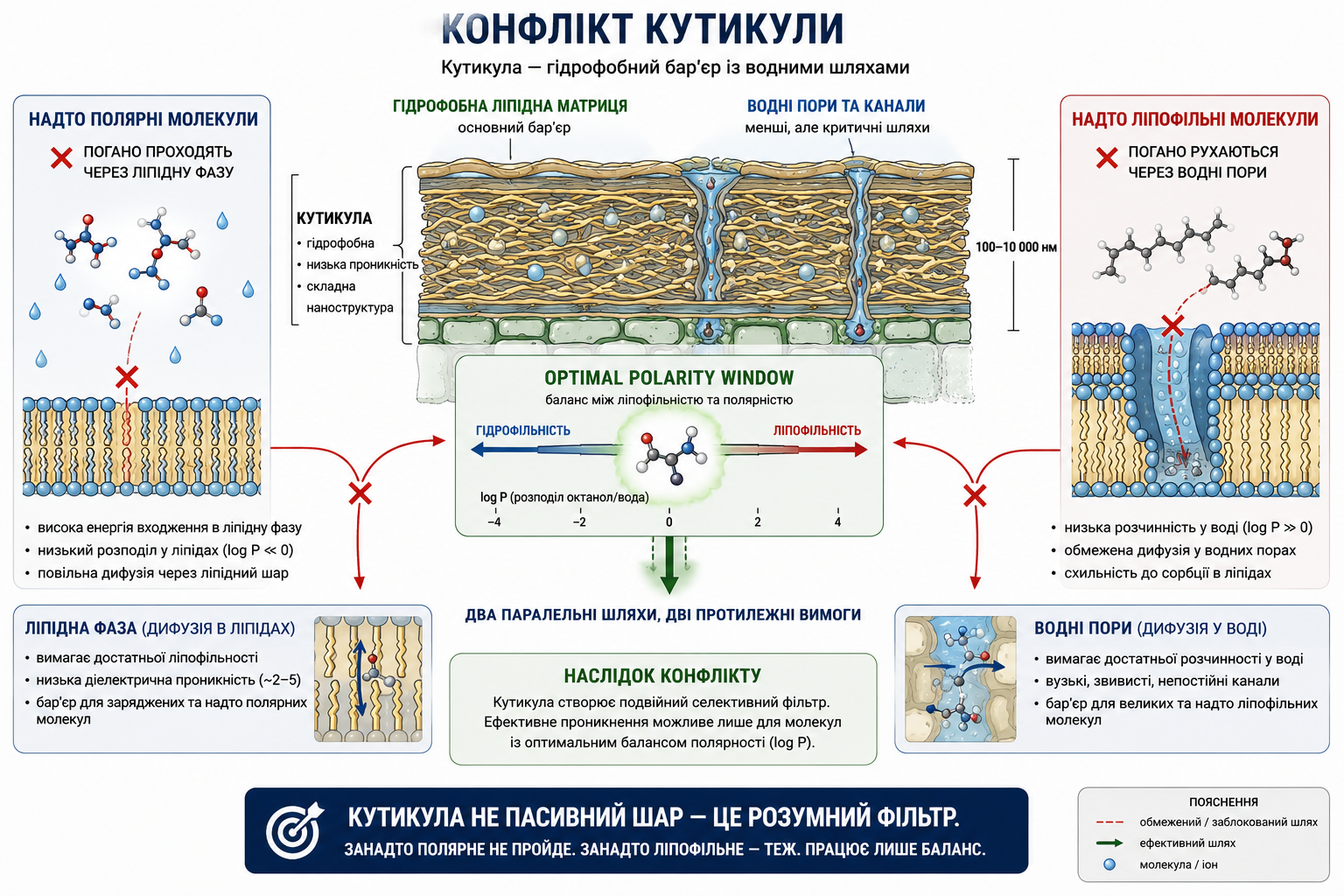
Мал. 7. Конфлікт полярності кутикули: фізико-хімічний компроміс між ліпофільністю та гідрофільністю при позакореневому поглинанні. Схематичне зображення подвійної селективної природи рослинної кутикули як бар’єра для позакореневого проникнення мікроелементів і комплексів. Кутикула одночасно містить гідрофобну ліпідну матрицю та водні пори, що створює фундаментальний фізико-хімічний конфлікт для транспорту молекул. Надто полярні або заряджені сполуки мають обмежену дифузію через ліпідну фазу, тоді як надто ліпофільні молекули погано переміщуються через водні канали та характеризуються низькою розчинністю у водному середовищі. Ефективне проникнення можливе лише в межах “оптимального вікна полярності”, де досягається баланс між гідрофільністю та ліпофільністю. Рисунок ілюструє, що проникнення через кутикулу визначається не маркетинговою назвою формуляції, а фундаментальними законами дифузії, полярності та фазового розподілу.
ПОЛІОЛИ І “НАНО” — НАЙБІЛЬШИЙ МАРКЕТИНГОВИЙ ФІКЦІОН.
Поліоли, цукрові спирти та “нано-форми” стали одними з найсильніших маркетингових символів сучасного ринку мікродобрив і біостимуляторів. Саме навколо манітолу, сорбітолу, різних цукрів та нано-структур сьогодні будується значна частина рекламних концепцій про “революційне проникнення”, “біологічний транспорт” і “максимальну біодоступність”. У презентаціях ці речовини часто описують як майже інтелектуальні системи доставки, здатні переносити мікроелементи через кутикулу прямо в клітину рослини. Формується враження, що достатньо зв’язати елемент із поліолом або перевести його в “нано-форму” — і проблема засвоєння автоматично зникає. Але саме тут і виникає один із найбільших розривів між маркетинговою риторикою та реальною фізіологією рослини.
Поліоли — манітол, сорбітол та інші багатоатомні спирти — дійсно можуть утворювати комплекси з мікроелементами та впливати на властивості розчину. Проте їхня реальна роль значно прозаїчніша, ніж це подає реклама. Вони не “відкривають” кутикулу, не створюють нових мембранних транспортерів і не змінюють базову архітектуру клітинного транспорту. Рослина не має спеціальних механізмів, які б автоматично розпізнавали поліольний комплекс як “пропуск” для проникнення в клітину. Усі фундаментальні фізіологічні бар’єри залишаються незмінними: кутикула продовжує бути гідрофобним захисним шаром, клітинна стінка — структурним бар’єром, а мембрани — селективною системою з білковими транспортерами, які працюють лише з певними іонами або молекулами.
У багатьох випадках поліоли виконують значно простіші фізико-хімічні функції. Вони можуть покращувати стабільність розчину, знижувати ризик кристалізації солей, впливати на в’язкість краплі або покращувати змочування листкової поверхні. Також вони можуть підвищувати сумісність компонентів у баковій суміші та частково впливати на швидкість висихання робочого розчину. Усе це важливі технологічні властивості, але вони не є “революцією засвоєння”. Це не переписування фізіології рослини, а лише корекція фізики розчину.
Саме тут маркетинг часто підміняє поняття. Покращення поведінки краплі на поверхні листка починають подавати як доказ “покращеного проникнення в клітину”. Але контакт із листком не дорівнює транспорту через мембрану. Те, що розчин довше залишається в рідкому стані або краще розтікається по кутикулі, ще не означає автоматичного включення елемента в метаболізм рослини. У реальності між цими етапами існує складна система бар’єрів і обмежень.
Подібна ситуація виникає і з “нано-формулами”. Сам термін “нано” в агрохімії часто використовується як маркетинговий символ високої технологічності, хоча далеко не завжди має чітке фізико-хімічне обґрунтування. У рекламі створюється ілюзія, що зменшення розміру частинок автоматично забезпечує проникнення через кутикулу та мембрани. Але реальна біофізика рослини набагато складніша. Наночастинка не отримує “пропуск” у клітину лише через свої розміри. Вона все одно повинна пройти через гідрофобні шари кутикули, пористу структуру клітинної стінки та селективну мембранну систему. Більше того, багато наночастинок у реальних умовах можуть агломеруватися, змінювати заряд, взаємодіяти з компонентами поверхні листка або втрачати свої властивості ще до проникнення.
Найважливіше те, що ні поліоли, ні “нано-форми” не усувають фундаментального фізіологічного конфлікту системи: рослина контролює транспорт речовин через мембрани вибірково, а не за рекламною назвою препарату. Більшість мікроелементів усе одно повинні перейти у відповідну іонну форму, яку можуть розпізнати транспортні білки клітини. Саме тому навіть найскладніший “біоактивний комплекс” часто в кінцевому підсумку працює через ті ж самі механізми, що й класичні форми мікроелементів.
У підсумку поліоли, цукрові спирти та “нано-технології” в агрохімії найчастіше не є революцією у біології рослини. Вони можуть покращувати окремі фізико-хімічні параметри робочого розчину — стабільність, змочування, сумісність або поведінку краплі. Але це не означає створення нових механізмів засвоєння. Тому головна проблема полягає не в самих поліолах чи нано-формах, а в тому, що індустрія часто продає покращення фізики краплі як революцію клітинної фізіології, хоча між цими речами існує принципова різниця.
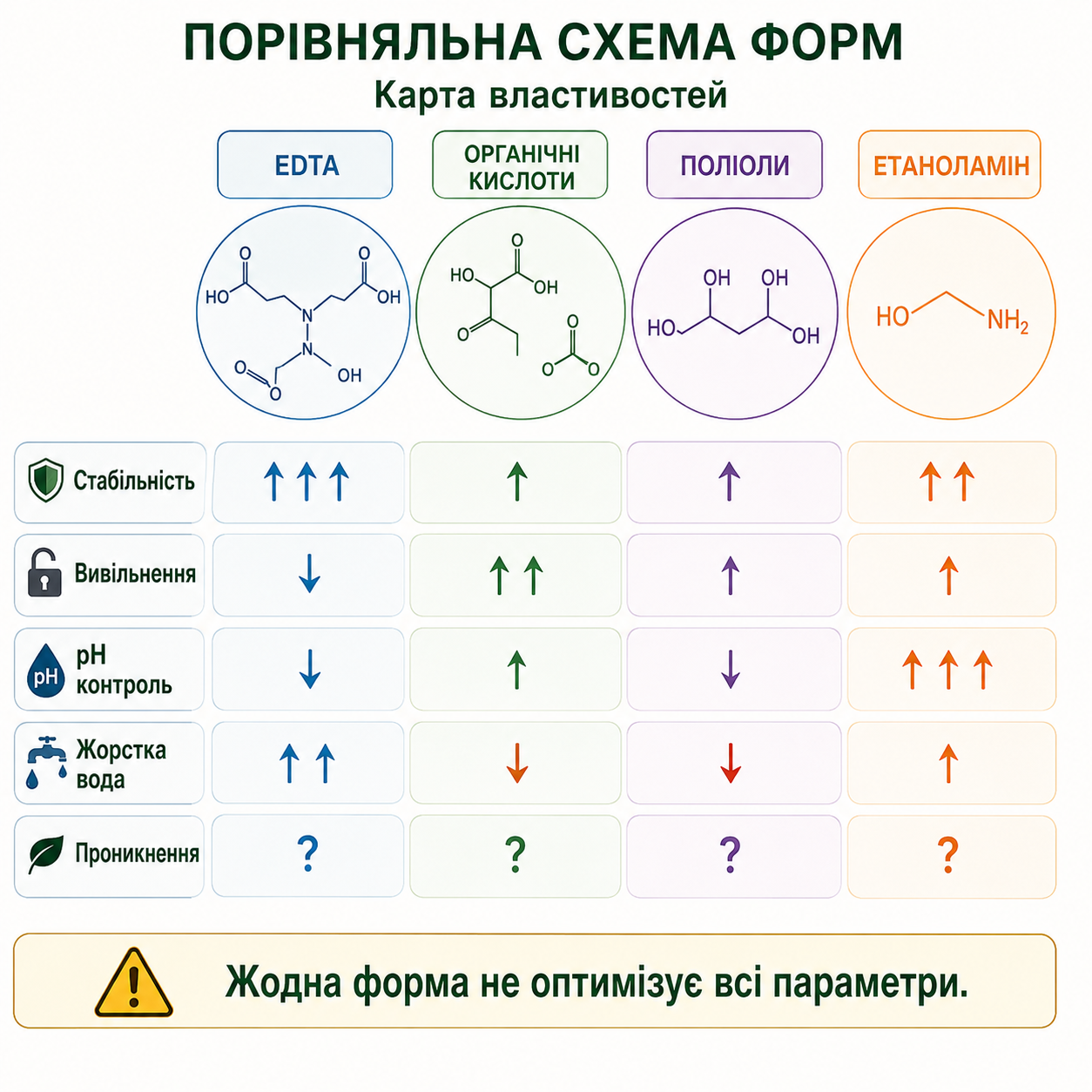
Мал. 8. Порівняльна схема властивостей хелатних і комплексоутворюючих форм мікроелементів. На рисунку представлено порівняльну карту властивостей різних форм комплексоутворювачів і лігандів, що використовуються у спеціальних добривах та мікроелементному живленні рослин: EDTA, органічних кислот, поліолів та етаноламіну. Схема демонструє відмінності між формами за стабільністю комплексів, швидкістю вивільнення елементів, здатністю контролювати pH, сумісністю з жорсткою водою та потенційним проникненням через бар’єри рослинної поверхні. Показано, що жодна форма не забезпечує одночасної оптимізації всіх фізико-хімічних і біологічних параметрів.
ЕТАНОЛАМІН І “СУЧАСНІ ХЕЛАТИ” — ПРОМІЖНА СІРА ЗОНА.
Етаноламін у мікродобривах та так званих “сучасних хелатах” займає дуже цікаву проміжну позицію між класичною координаційною хімією та маркетинговими інтерпретаціями “нового покоління” агрохімії. У рекламних матеріалах його часто подають як “інтелектуальний ліганд”, “біологічно сумісний носій”, “органічний активатор засвоєння” або навіть “альтернативу EDTA”. Проте з точки зору фундаментальної хімії етаноламін не є повноцінним хелатуючим агентом у класичному розумінні, яким є, наприклад, EDTA, DTPA чи EDDHA. Його роль значно скромніша і значно ближча до регуляції середовища, часткової комплексоутворюючої стабілізації та підтримки розчинності металів у робочому розчині.
Класичний хелат — це структура, яка утворює з іоном металу стабільне циклічне координаційне кільце. Саме багатозубість ліганду забезпечує високу термодинамічну стабільність комплексу та захист іона від осадження, гідролізу чи переходу в нерозчинні форми. EDTA або DTPA буквально “обгортають” метал, формуючи дуже стабільні комплекси. Етаноламін працює зовсім інакше. Його молекула має лише одну аміногрупу та одну гідроксильну групу, тому його здатність до координації металів обмежена. Він може тимчасово взаємодіяти з катіонами, частково стабілізувати їх у розчині та змінювати локальне хімічне середовище, але не створює настільки міцних координаційних структур, як класичні хелатори.
Саме тому етаноламінові комплекси часто мають слабшу стабільність, особливо при коливаннях pH, жорсткій воді або високій концентрації фосфатів і карбонатів. У певних умовах метал може легко вивільнятися з такого комплексу та переходити у нерозчинні форми. Однак ця ж “слабкість” інколи розглядається як перевага. Надто стабільні хелати іноді утримують метал настільки міцно, що його доступність для рослини знижується або потребує додаткових фізіологічних механізмів для вивільнення. Етаноламінові системи створюють компроміс: метал достатньо стабілізований для збереження розчинності, але не настільки жорстко зв’язаний, як у випадку EDTA або EDDHA.
Крім того, етаноламін виконує дуже важливу роль регулятора pH. Це одна з його ключових функцій у рідких добривах. Він дозволяє утримувати метал у більш стабільному стані за рахунок зміни кислотно-лужного середовища, зменшуючи ризик швидкого випадіння осаду. Часто саме контроль pH, а не власне “хелатування”, пояснює покращення стабільності препарату. Тобто реальний механізм дії значно прозаїчніший, ніж це подається в рекламі “біо-хелатів нового покоління”.
Саме тут і виникає так звана “сіра зона” сучасного маркетингу агрохімії. Багато продуктів, які називають “новітніми органічними хелатами”, “біохелатами” або “амінохелатами”, фактично є сумішами слабких комплексоутворювачів, буферів і розчинників, де етаноламін виконує допоміжну функцію. З наукової точки зору це не революція у живленні рослин, а лише інший баланс між стабільністю, розчинністю, ціною та технологічністю препарату. Але маркетингово такі продукти часто подаються як принципово новий клас речовин, що нібито кардинально перевершує класичні хелати.
Особливо поширеним є твердження: “хелат = 100% засвоєння”. Це один із найбільш стійких міфів у ринку мікродобрив. У свідомості багатьох агрономів слово “хелат” автоматично асоціюється з максимальною ефективністю та гарантованим поглинанням елементу рослиною. Насправді хелатування вирішує лише одну проблему — підтримання металу у доступному та стабільному стані в розчині. Воно не гарантує проникнення через кутикулу листка, не забезпечує транспорту через мембрани, не усуває фізіологічні обмеження рослини та не визначає ефективність внутрішньоклітинного метаболізму.
Поглинання елементів живлення визначається насамперед фізіологією рослини: активністю транспортерів, енергетичним станом клітини, температурою, водним режимом, функціонуванням продихів, станом кутикули, рівнем стресу, швидкістю транспірації та потребами метаболізму. Якщо рослина перебуває у стані стресу, має пошкоджену кореневу систему або низьку активність мембранного транспорту, навіть ідеально стабільний хелат не забезпечить високого засвоєння. У багатьох випадках саме фізіологічний стан культури є головним лімітуючим фактором, а не форма елементу.
Більше того, іноді надмірна стабільність хелату навіть уповільнює доступність металу. Рослина повинна або руйнувати комплекс, або використовувати спеціалізовані механізми поглинання. Тому “сильніший хелат” не завжди означає “краще живлення”. У реальних польових умовах ефективність часто визначається не абсолютною стабільністю комплексу, а правильним балансом між стабільністю, мобільністю та швидкістю вивільнення іона.
Таким чином, етаноламінові системи — це не “магічна нова хімія”, а скоріше технологічний компроміс. Вони можуть бути корисними у певних формулах, особливо для рідких добрив, позакореневого живлення або сумісності бакових сумішей. Але їх не можна розглядати як універсальну революційну заміну класичних хелатів. І головне — сама наявність слова “хелат” у назві продукту не означає автоматично високого засвоєння, оскільки реальна ефективність живлення визначається не маркетинговим терміном, а складною взаємодією хімії розчину та фізіології рослини.
ГОЛОВНА ПРОБЛЕМА ІНДУСТРІЇ: ПІДМІНА ПОНЯТЬ.
Головна проблема сучасного агробізнесу полягає не стільки у відсутності ефективних рішень, скільки у способі їх подачі та інтерпретації. Індустрія дедалі рідше продає просто ефективність у науковому розумінні цього слова — тобто прогнозовану, відтворювану дію речовини в конкретних умовах. Натомість вона продає складність замість зрозумілості, терміни замість механізмів і презентації замість реальних експериментальних доказів. У результаті формується паралельна реальність, де рівень “сучасності” продукту визначається не його фізико-хімічними властивостями, а насиченістю маркетингового словника.
Терміни на кшталт “нано”, “біо”, “інтелектуальний”, “активований”, “фізіологічно адаптований” або “молекулярно керований” не є науковими категоріями у строгому сенсі. Вони не описують конкретні параметри координаційної хімії, кінетику реакцій, стабільність комплексів або механізми транспорту через мембрани. Це радше маркетингові етикетки, наклеєні на добре відомі класи сполук — хелати, солі, комплекси з органічними кислотами, поліолами або амінними похідними. Проте завдяки складній термінології створюється враження принципово нової технології, хоча базова хімічна сутність часто залишається тією самою.
Ключова помилка маркетингу полягає в системній підміні логіки. Замість того щоб описувати багатофакторну систему взаємодії “розчин — листок — клітина — метаболізм”, вона зводиться до одного простого меседжу: “це нова форма — значить вона краща”. Така логіка виглядає переконливо з точки зору продажів, але вона повністю ігнорує фізико-хімічну реальність. У реальних біологічних системах не існує лінійного прогресу, де кожне нове покоління форм автоматично перевершує попереднє.
Насправді засвоєння мікроелементів є системою обмежень, а не системою безмежної оптимізації. Не існує універсально “кращої” форми мікроелемента, яка однаково ефективно працює в усіх умовах. Будь-яка хімічна форма — це лише компроміс між кількома взаємно конфліктними параметрами: стабільністю в розчині, стійкістю до антагоністів, швидкістю дисоціації, здатністю до проникнення через кутикулу та можливістю включення в метаболічні процеси всередині клітини.
Якщо посилюється стабільність комплексу — зростає контроль у баковій суміші, але зменшується швидкість вивільнення іону в зоні поглинання. Якщо підвищується реактивність і легкість дисоціації — покращується доступність, але падає стійкість у складних фізико-хімічних умовах. Якщо оптимізується проникнення — це часто відбувається за рахунок втрати контролю над транспортом і стабільністю системи. Тобто кожне покращення одного параметра неминуче створює обмеження в іншому.
Саме цей фундаментальний баланс майже ніколи не пояснюється в маркетингових матеріалах. Замість опису компромісів і умов ефективності споживачу пропонується спрощена модель, де нова формула нібито одночасно покращує всі етапи процесу: і стабілізує, і транспортує, і проникає, і повністю засвоюється. Але така система суперечить базовим принципам координаційної хімії та фізіології рослин.
У реальній агрохімії питання не може звучати як “яка форма найкраща”, тому що універсально найкращої форми не існує. Коректне формулювання звучить інакше: яка форма є оптимальною для конкретного набору умов — типу ґрунту або води, культури, фази розвитку рослини, температури, вологості та способу внесення. Саме тут і виникає головний розрив між науковою логікою і маркетинговою: наука оперує системою обмежень і умов, тоді як маркетинг — системою універсальних обіцянок.
РЕАЛЬНІСТЬ, ЯКУ НЕ ПОКАЗУЮТЬ У РЕКЛАМІ.
Реальність польового застосування мікродобрив і позакореневого живлення набагато складніша, ніж це показують рекламні матеріали виробників. У маркетингу основний акцент майже завжди робиться на “унікальній формі”, “інноваційному носії”, “нано-технології”, “біокомплексі” або “сучасному хелаті”. Створюється враження, що саме назва технології автоматично гарантує високу ефективність препарату. Але в реальному полі ситуація працює зовсім інакше. Для рослини не має принципового значення, як маркетологи назвали продукт. Фізіологія рослини не “читає” етикетку. Вона реагує лише на фізико-хімічні умови середовища та власний метаболічний стан.
Саме тому результат позакореневого підживлення визначається не красивими термінами, а конкретними польовими факторами. Одним із головних факторів є температура. Вона контролює швидкість випаровування робочого розчину, проникність кутикули, активність мембранного транспорту та інтенсивність метаболізму. При високих температурах крапля на листку може висохнути за кілька хвилин, не залишивши достатньо часу для проникнення елементів. Навіть найдорожчий “біохелат” у такій ситуації не покаже очікуваного ефекту, оскільки проблема не в формі елементу, а у фізиці висихання та обмеженні часу контакту.
Не менш важливу роль відіграє вологість повітря. При низькій вологості випаровування різко прискорюється, концентрація солей у краплі зростає, а ризик кристалізації на поверхні листка збільшується. У такому випадку значна частина діючої речовини може просто залишитися на поверхні у вигляді висохлого осаду. Навпаки, при помірній вологості крапля довше залишається рідкою, що суттєво покращує дифузію елементів через кутикулу. Саме тому інколи просте обприскування ввечері або рано вранці дає більший ефект, ніж заміна стандартного продукту на “ультрасучасний нанокомплекс”.
Критичне значення має і стан самого листка. Молодий листок, активно ростучий листок і старіючий листок мають абсолютно різні фізіологічні властивості. Товщина кутикули, кількість восків, інтенсивність транспірації, активність продихів і мембранних транспортерів суттєво відрізняються. У стресованої рослини кутикула часто ущільнюється, транспірація знижується, а транспорт через мембрани сповільнюється. У такому стані навіть ідеально стабілізований елемент не буде нормально поглинатися. Рослина повинна бути фізіологічно здатною прийняти і використати поживний елемент.
Дуже часто у рекламі майже не згадується швидкість висихання робочого розчину, хоча це один із ключових факторів ефективності позакореневого живлення. Фактично листок отримує лише той час, поки крапля залишається рідкою. Після висихання транспорт різко сповільнюється або взагалі припиняється. Саме тому ад’юванти, змочувачі та контроль умов внесення нерідко впливають на результат сильніше, ніж тип “хелату”. Інколи різниця між ранковим і денним внесенням може перевищувати різницю між двома “революційними” формуляціями.
Окремою недооціненою проблемою є тип води. Якість води для бакової суміші часто визначає стабільність препарату ще до контакту з рослиною. Жорстка вода з високим вмістом кальцію, магнію або бікарбонатів може частково руйнувати комплекси, викликати осадження або змінювати pH робочого розчину. Багато продуктів, які демонструють прекрасні результати у лабораторних умовах, у реальному господарстві втрачають ефективність саме через воду. Але це значно менш “продається”, ніж історії про “нанотехнології” чи “біоактивні комплекси”.
Ще важливішою є загальна фізіологія рослини. Якщо культура перебуває у водному стресі, дефіциті енергії, температурному шоку або фазі низької метаболічної активності, засвоєння буде обмеженим незалежно від форми препарату. Рослина — це не пасивна губка, яка автоматично поглинає все нанесене на листок. Поглинання — це активний фізіологічний процес, що потребує енергії, функціонуючих мембранних систем і відповідного метаболічного стану.
Саме тому в реальному полі вирішальними є не гучні маркетингові назви, а базові агрономічні фактори: коли внесли, при яких умовах внесли, як швидко висохла крапля, у якому стані була рослина, яка була вода, чи працював транспорт у тканинах, чи була рослина здатна засвоїти елемент. І дуже часто саме ці фактори пояснюють різницю між “препарат спрацював” і “препарат не дав ефекту”.
У підсумку реальна ефективність позакореневого живлення визначається не словами “нано-частинка”, “біокомплекс” або “сучасний хелат”. Вона визначається фізикою краплі, хімією розчину та фізіологією рослини. Саме ці три системи керують результатом у полі, тоді як маркетингові терміни дуже часто лише створюють ілюзію технологічної революції там, де насправді працюють давно відомі базові принципи агрохімії та рослинної фізіології.
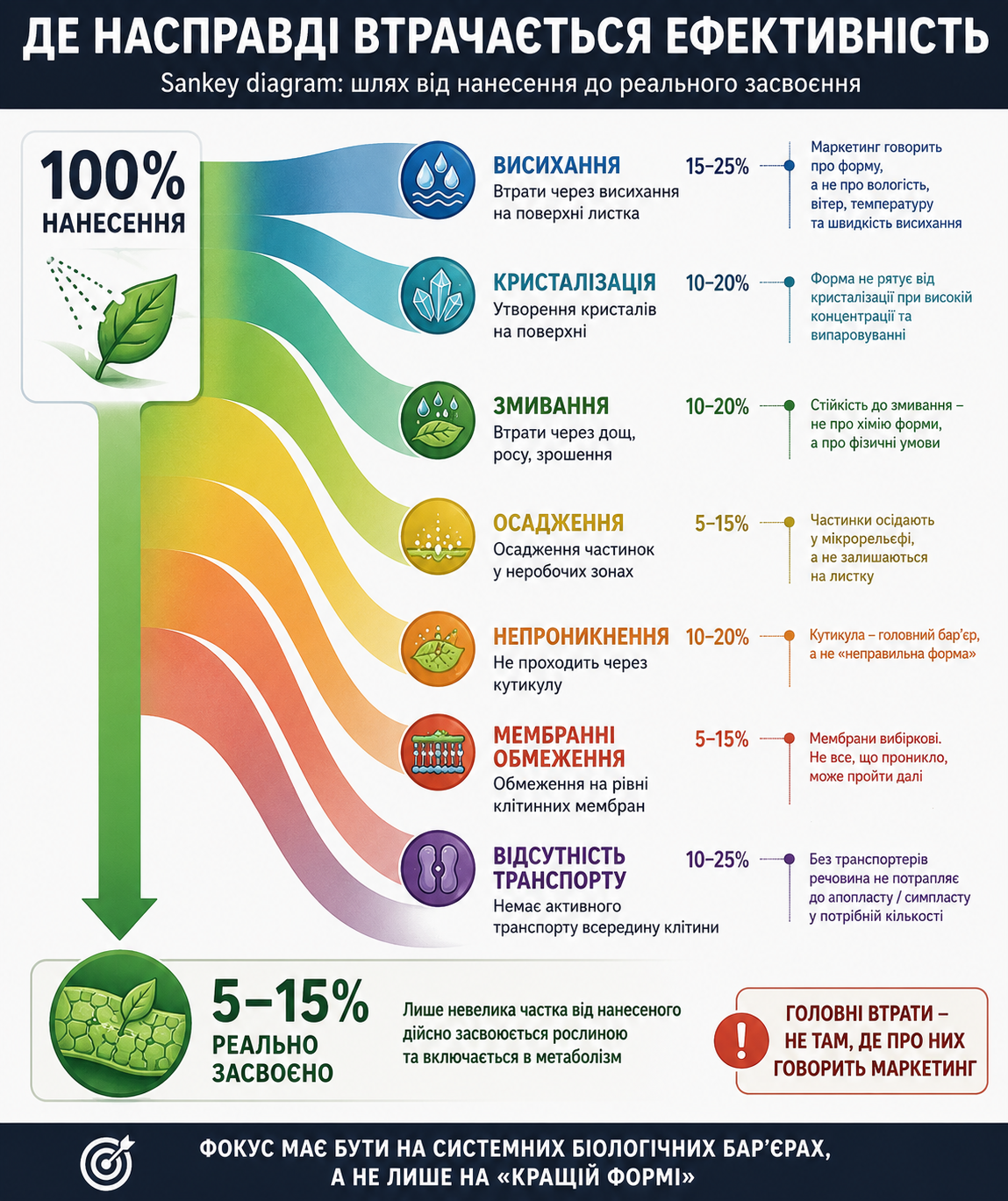
Мал. 9. Концептуальна схема втрати ефективності під час листкового внесення: ключові обмеження виникають переважно на етапах фізико-хімічної взаємодії з поверхнею та біологічного транспорту, а не лише через хімічну форму сполуки.
ФІНАЛ: ГОЛОВНА НЕЗРУЧНА ПРАВДА.
Фінальна і найменш зручна для індустрії правда полягає в тому, що значна частина сучасного аграрного ринку працює не як система створення принципово нових рішень, а як система переформатування вже давно відомої хімії під нові маркетингові оболонки. У більшості випадків мова йде не про появу нових механізмів дії або принципово інших фізіологічних шляхів засвоєння, а про зміну термінології, пакування та способу інтерпретації тих самих координаційних систем, які агрохімія використовує десятиліттями. Саме тому дуже часто складність назв і описів прямо пропорційна не реальній науковій новизні, а лише рівню маркетингової деталізації, за якою приховується звичайна, добре відома хімія.
Чим складніше і “технологічніше” звучить назва продукту – “нано-активований біокомплекс”, “інтелектуальний хелат”, “молекулярно адаптована система доставки” – тим менше шансів, що споживачу буде чітко і прозоро пояснено реальний механізм дії на рівні фізичної хімії та фізіології рослини. Замість опису стабільності комплексу, констант дисоціації, поведінки в жорсткій воді або реальних шляхів транспорту через кутикулу, використовується мова абстракцій, яка створює відчуття технологічного прориву, але не дає реального розуміння процесів.
Насправді ж базова структура системи залишається незмінною. У більшості випадків це все ті ж самі мікроелементи у вигляді іонів, солей або класичних комплексів, стабілізованих різними типами лігандів – від синтетичних хелатів до органічних кислот, поліолів або амінних похідних. Вони можуть відрізнятися за стабільністю, швидкістю вивільнення та поведінкою в різних середовищах, але вони не виходять за межі однієї і тієї ж фундаментальної хімічної системи координаційних взаємодій.
Саме тому ключова правда полягає в тому, що не існує “магічних форм” мікроелементів, які автоматично вирішують проблему живлення рослин незалежно від умов. Не існує універсальної молекули, яка одночасно ідеально стабільна в баковій суміші, миттєво проникає через кутикулу і повністю контролюється всередині клітини. Будь-яка форма – це лише різний набір компромісів між стабільністю, мобільністю, швидкістю дисоціації та біодоступністю.
І саме тому реальна ефективність ніколи не визначається лише назвою продукту або його “поколінням”. Вона завжди залежить від конкретних умов: властивостей води і ґрунту, температури, вологості, фізіологічного стану рослини, фази розвитку та способу внесення. Те, що в одних умовах працює добре, в інших може бути нейтральним або навіть менш ефективним, незалежно від того, наскільки “сучасною” є формула.
У підсумку залишається проста, але незручна для маркетингу істина: агрохімія – це не ринок універсальних інновацій, а система фізико-хімічних компромісів, де результат завжди визначається умовами середовища і фізіологією рослини, а не красивістю термінів у презентації.
ПРО ЕКОНОМІКУ МАРКЕТИНГУ.
Окремою причиною стрімкого поширення “інноваційних” форм мікроелементів, нанодобрив, біокомплексів та складних хелатних систем є не лише хімія чи фізіологія рослин, а економіка самого аграрного ринку. Важливо розуміти, що більшість подібних тенденцій виникають не через “змову” виробників, а через цілком логічні ринкові механізми. Сучасний ринок добрив — це висококонкурентне середовище, де класичні мінеральні солі давно стали стандартизованим товаром із низькою маржинальністю. Сульфат цинку, сульфат марганцю, борна кислота або молібдат амонію мають відносно простий хімічний склад, добре відомі механізми дії та легко порівнюються між різними виробниками. Через це конкуренція між такими продуктами переважно переходить у площину ціни, а прибутковість для виробника зменшується.
Натомість складні “нано-біокомплекси”, “органо-мінеральні матриці”, “біоактивовані хелати”, “супрамолекулярні системи доставки” або інші технологічно звучні форми дозволяють створити принципово інший тип ринкового продукту. По-перше, складна назва формує диференціацію. Якщо звичайний сульфат цинку є зрозумілим і легко порівнюваним товаром, то “наноактивований поліфункціональний цинковий біокомплекс” практично неможливо безпосередньо співставити з аналогами. Це створює інформаційну асиметрію між виробником і покупцем, де фермер часто не має інструментів для незалежної перевірки реального механізму дії або об’єктивної оцінки заявленої “унікальності”.
По-друге, складний продукт значно легше патентувати та брендувати. Класичні солі або прості хелати важко зробити ексклюзивними, тоді як специфічна комбінація лігандів, допоміжних компонентів, “наноструктур” або технологічних назв створює ефект унікальної технології. Це дозволяє компанії формувати вищу додану вартість навіть у випадках, коли базова фізіологічна функція препарату залишається тією самою — доставка іона мікроелемента до рослини.
Особливу роль відіграє те, що в системі живлення рослин дуже складно швидко й однозначно перевірити причинно-наслідковий зв’язок між препаратом і кінцевим результатом. Урожайність визначається десятками факторів: погодою, ґрунтом, вологою, гібридом, густотою стояння, фітосанітарним станом, живленням макроелементами, стресами та технологією вирощування. На цьому фоні навіть випадкові коливання врожайності можуть інтерпретуватися як доказ ефективності певного “інноваційного” продукту. Якщо фермер отримав додаткові 0,2–0,4 т/га після внесення складного препарату, практично неможливо без багаторазових контрольованих дослідів точно визначити, яка частина цього ефекту пов’язана саме з хімічною формою мікроелемента, а яка — з погодою чи природною варіабельністю поля.
Саме тому маркетинг у сфері мікродобрив часто продає не стільки безпосередньо вимірюваний ефект, скільки очікування технологічної переваги. Це дуже характерно для продуктів, де механізм дії є невидимим для кінцевого користувача. Фермер не може візуально оцінити, у якій саме формі цинк проходить через кутикулу, чи зберігається “наноструктура” після висихання краплі, або який відсоток комплексу реально досягає транспортних систем клітини. У результаті маркетингова цінність складної термінології різко зростає. Такі поняття, як “нанотехнологія”, “біоактивація”, “квантова стабілізація”, “інтелектуальна доставка” або “молекулярне проникнення”, часто працюють як інструменти формування сприйняття інноваційності навіть тоді, коли реальні фізіологічні механізми залишаються класичними.
Важливо також розуміти, що аграрний ринок психологічно дуже чутливий до обіцянок підвищення ефективності. Умови сучасного землеробства стають дедалі стресовішими: нестабільний клімат, посухи, високі ціни на добрива, деградація ґрунтів і зростання ризиків виробництва створюють постійний попит на “технологічне рішення”, яке здатне забезпечити додаткову стабільність урожаю. На цьому фоні продукти, що позиціонуються як “революційні” або “принципово нові”, отримують значну комерційну перевагу навіть без радикально нового механізму дії.
При цьому сама наявність маркетингової складової не означає автоматично, що продукт є неефективним або шахрайським. Частина сучасних комплексних форм дійсно може мати певні технологічні переваги: кращу стабільність у бакових сумішах, меншу фітотоксичність, вищу сумісність із засобами захисту, покращене утримання на листку або повільніше вивільнення елемента. Проте економічна логіка ринку стимулює постійне перебільшення ступеня цієї “унікальності”. Чим складніше пояснити механізм препарату, тим легше сформувати навколо нього образ високотехнологічного рішення з високою доданою вартістю.
У результаті сучасний ринок мікродобрив значною мірою функціонує не лише як ринок агрохімії, а й як ринок інформації, довіри та технологічного позиціонування. Саме тому критично важливо відокремлювати реальні фізіологічні переваги препарату від маркетингової надбудови навколо нього. Для об’єктивної оцінки продукту недостатньо лише складної назви або заяв про “нано-” чи “біоактивність”; необхідні польові дані, повторювані результати, зрозумілий механізм дії та економічна оцінка того, чи виправдовує потенційний ефект суттєво вищу вартість препарату порівняно з класичними формами елементів.
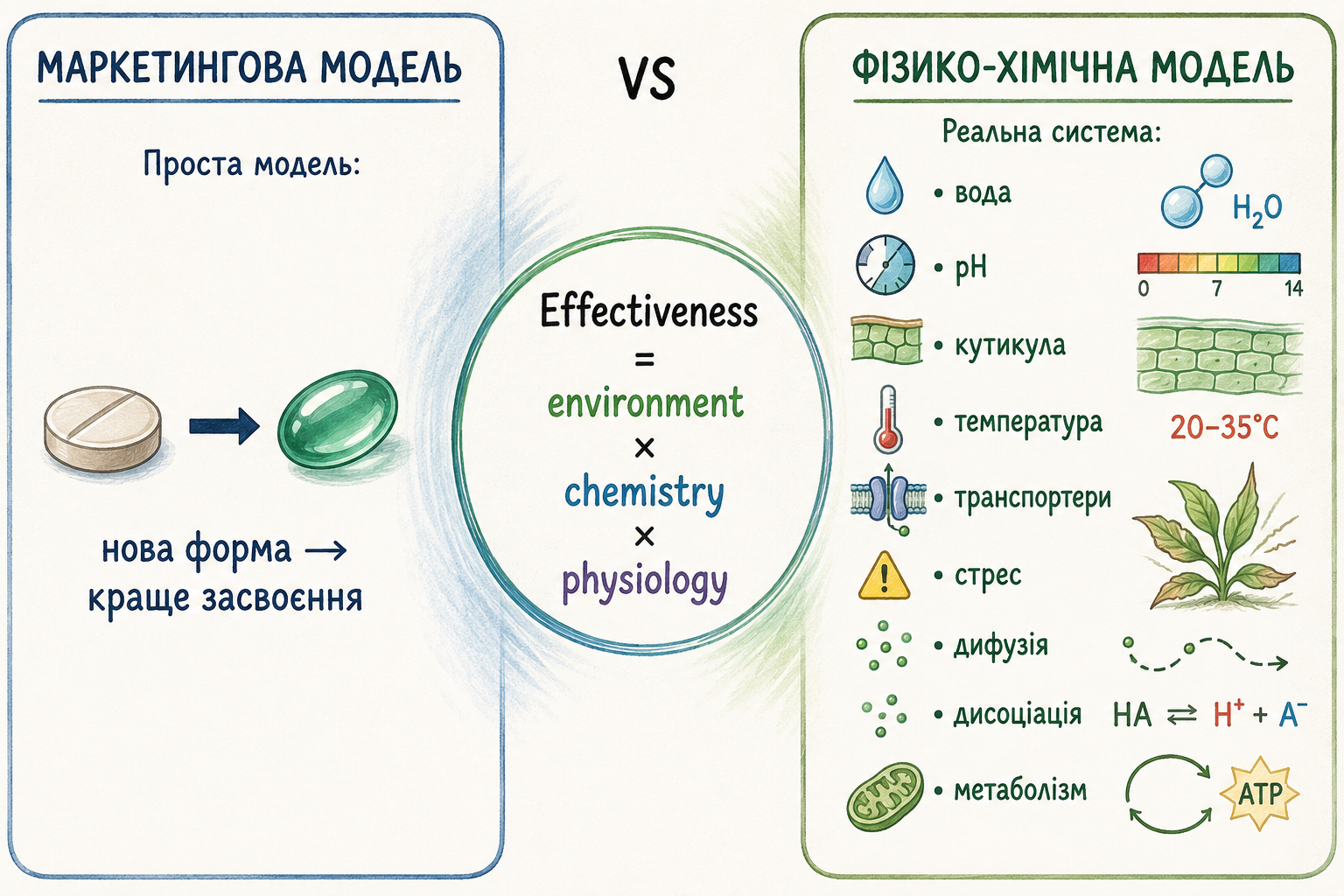
Мал. 10. Маркетингова модель vs фізико-хімічна модель ефективності засвоєння. Порівняння спрощеної маркетингової моделі («нова форма → краще засвоєння») з фізико-хімічною моделлю, у якій ефективність визначається взаємодією середовища, хімічних властивостей речовини та фізіологічних процесів.
ВИСНОВКИ
У сучасній агрохімії одна з найбільш недооцінених, але критичних проблем полягає в тому, що різні форми мікроелементів – класичні хелати (EDTA, DTPA, EDDHA), комплекси з органічними кислотами, поліолами (манітол, сорбітол), цукрами та етаноламіном – у маркетингу подаються як “покращені версії” однієї і тієї ж технології живлення. Створюється ілюзія, що існує лінійний прогрес: від простих солей до “сучасних нанокомплексів”, які нібито одночасно стабільніші, швидше засвоюються, краще проникають у листок і повністю вирішують проблему живлення рослин. Насправді ж це не еволюційна шкала, а набір різних хімічних інструментів, які працюють на різних етапах одного складного процесу.
Фізіологія засвоєння мікроелементів – це не один механізм, а багатоступенева система: стабілізація елемента в розчині, транспортування до поверхні листка або кореня, проходження бар’єрів (кутикула, клітинна стінка, мембрани) і лише потім внутрішньоклітинне включення в метаболізм. Кожен із цих етапів має свої фізико-хімічні обмеження, і жодна форма не може одночасно бути і максимально стабільною в баку, і максимально швидкою в вивільненні, і максимально проникною в тканини рослини.
Саме тут виникає ключова помилка маркетингу – підміна функцій. Наприклад, класичні хелати EDTA або DTPA справді ефективні на етапі стабілізації: вони утримують іон металу в розчині, запобігають осадженню і захищають від антагонізму з кальцієм, магнієм чи карбонатами. Але їхня висока стабільність може обмежувати швидкість вивільнення іону в зоні поглинання. Навпаки, комплекси з органічними кислотами швидше віддають іон, але слабше захищають його в жорсткій воді або складних бакових сумішах.
Поліоли (манітол, сорбітол) і цукри займають окрему нішу. Їх часто позиціонують як “біоміметичні транспортери” або навіть структури, що нібито покращують проникнення в клітину. Але в реальності їх роль значно простіша: стабілізація розчину, покращення сумісності компонентів і зміна фізичних властивостей краплі. Вони можуть впливати на змочування листка, але не створюють “магічного транспорту” через кутикулу.
Етаноламінні комплекси також часто подають як сучасну альтернативу хелатам. Насправді це проміжна система: етаноламін виконує роль слабкого ліганду і регулятора pH, але не забезпечує тієї передбачуваної стабільності, як класичні хелатуючі агенти. Їх поведінка сильно залежить від умов середовища – pH, температури та іонного складу води.
Якщо розглядати всю систему цілісно, стає очевидно: кожен тип комплексу оптимізує лише одну ділянку ланцюга “стабілізація → транспортування → вивільнення → засвоєння”. І головний компроміс полягає в тому, що посилення одного етапу майже завжди погіршує інший. Чим стабільніший комплекс – тим гірше він віддає іон. Чим швидше він дисоціює — тим гірше він захищений у баковій суміші.
Саме тому в реальній агрохімії питання не стоїть як “що краще – хелати чи поліоли”, бо це хибна постановка задачі. Правильне питання інше: для якого середовища, для якої культури, на якій фазі розвитку і для якого етапу живлення обрана ця форма. Жорстка вода, температура, вологість, тип кутикули, швидкість випаровування і фізіологічний стан рослини часто мають значно більший вплив, ніж сама форма комплексу.
Таким чином, головний розрив між маркетингом і наукою полягає в заміні багатофакторної фізико-хімічної системи простим гаслом “нова форма = краща ефективність”. У реальності ж кожна форма мікроелемента – це не універсальне покращення, а лише компроміс між стабільністю, мобільністю та біодоступністю, який працює тільки в конкретних умовах і ніколи не є абсолютним.
У більшості польових умов обмеженням є не тип комплексу мікроелемента, а зовсім інші фактори: швидкість висихання краплі, температура, вологість повітря, стан кутикули, рівень транспірації та фізіологічний стрес рослини. Навіть ідеально підібраний хелат або “нано-комплекс” не може компенсувати ситуацію, коли листок закритий через посуху або коли рослина фізіологічно не здатна транспортувати елементи через знижену активність флоеми.
Більшість заяв про “покращене засвоєння” базуються не на прямому вимірюванні проникнення іонів у клітину, а на непрямих польових ефектах – приріст маси, зелений колір, візуальна реакція рослини. Але ці параметри не дозволяють розділити ефект живлення, ефект ПАР, ефект води або загального зменшення стресу. У результаті будь-яка комплексна органічна форма може виглядати “ефективнішою” навіть без доведення реального шляху іону в тканини рослини.
Важливо також, що ефективність мікроелементів сильно залежить від способу внесення. Те, що працює в ґрунті, не обов’язково працює через листок. У ґрунті домінують процеси дифузії, адсорбції та іонного обміну, тоді як у листковому живленні ключовими є змочування, проникнення через кутикулу та локальна дисоціація. Тому одна й та сама форма комплексу може поводитися кардинально по-різному залежно від того, де вона застосовується.
Чим більше технологія оптимізує один параметр – тим більше вона обмежує інший. Підвищення стабільності зменшує швидкість вивільнення. Підвищення проникності часто знижує захист у баковій суміші. Підвищення розчинності може зменшувати контрольованість доставки іону в потрібну тканину. У результаті не існує універсально “кращої” форми – існує лише баланс властивостей під конкретну задачу.
У більшості випадків фермер купує не кращу хімію, а іншу форму того самого компромісу.
Хаблак Сергій, д.б.н., агроном-експерт, агроаудитор, 066-44-266-08
Читайте нас у TelegramПов’язані теми:









Посилюється фітосанітарний контроль при експорті пшениці до Турецької Республіки
03.08.2026, 10:15
Новини

Перший у штаті Новий Південний Уельс завод зеленого аміаку буде використовувати сонячну енергетику
03.08.2026, 08:22
Добрива

Нескладно відрізнити коника від сарани, хоч вони й схожі
02.08.2026, 19:03
Новини

«Укрзалізниця» з 31 липня проіндексувала тарифи на вантажні перевезення на 30%
01.08.2026, 22:36
Агропромисловість