
Концепція «Імунно-метаболічного управління культурою»
25.12.2025
Стан імунітету рослин у світі та чому ринок потребує нових антистресових технологій

Мал. 1 Абіотичні фактори знижують урожайності більше ніж на 60%.
Вступ.
За останні два десятиліття світове агровиробництво увійшло в період безпрецедентної нестабільності. Кліматичні зміни трансформували агросистеми з відносно прогнозованих у високоризикові, де поєднуються температурні аномалії, дефіцит або надлишок вологи, деградація ґрунтів, порушення мікробіоти та різке зростання інфекційного тиску. Рослина дедалі частіше перебуває не в умовах одного домінуючого стресу, а під впливом комплексного, хронічного та накладеного стресового навантаження.
Одночасно з абіотичними факторами спостерігається швидка еволюція патогенів. Нові раси грибів, бактерій і вірусів, а також зміна їхньої агресивності та екологічної ніші, знижують ефективність традиційних систем захисту. Навіть високопродуктивні сорти з заявленою генетичною стійкістю дедалі частіше демонструють втрату адаптивного потенціалу вже на ранніх етапах онтогенезу.
Попри значний прогрес у селекції, агрохімії та біотехнологіях, підходи до управління стійкістю рослин довгий час залишалися фрагментарними. Селекційні програми переважно орієнтувалися на окремі гени резистентності або продуктивні ознаки, тоді як агрохімічні рішення фокусувалися на усуненні окремих факторів стресу — патогена, дефіциту елементів живлення чи водного дисбалансу. У результаті ринок отримував локальні інструменти, які не забезпечували стабільного ефекту в умовах комбінованого стресу.
Відсутність цілісної, інтегрованої теорії рослинного імунітету призвела до того, що більшість антистресових препаратів працюють емпірично: їхня дія часто непередбачувана, залежна від фази розвитку культури, погодних умов або технологічного фону. Це формує розрив між очікуваннями виробника і реальним польовим результатом, знижуючи довіру до інноваційних продуктів.
Водночас сучасна наука накопичила значний обсяг даних про молекулярну організацію імунітету рослин — від рецепторів розпізнавання стресових сигналів до складних мереж гормональної та метаболічної регуляції. Стає очевидно, що стійкість рослини — це не окрема властивість, а динамічний системний стан, який формується взаємодією генетики, фізіології, сигналінгу та середовища.
У цих умовах аграрний ринок об’єктивно потребує нового покоління антистресових технологій — не просто стимуляторів росту чи коректорів живлення, а інструментів, здатних керувати імунним статусом рослини на рівні регуляторних механізмів. Йдеться про підходи, засновані на молекулярній логіці адаптації: активації неспецифічної резистентності, підвищенні толерантності до комплексного стресу та збереженні продуктивності без надмірного навантаження на агроекосистему.
Саме в цьому контексті формуються нові наукові моделі і практичні рішення, що змінюють уявлення про рослинний імунітет — від реактивного захисту до проактивного управління стійкістю. Їхній аналіз є критично важливим для розуміння того, куди рухається світова агроіндустрія і чому інвестиції в інтелектуальні антистресові технології стають не опцією, а необхідністю.
Глобальна ситуація: головні виклики для продуктивності рослин.
У сучасному агровиробництві ключовим обмежувальним фактором продуктивності дедалі більше стають не окремі хвороби чи шкідники, а сукупний тиск абіотичних стресів. За узагальненими оцінками міжнародних аналітичних і наукових джерел, абіотичні фактори відповідають за 60–70% глобальних втрат урожайності, тоді як частка біотичних чинників становить близько 30–40%. Такий розподіл принципово змінює логіку управління ризиками в агросистемах і демонструє, що основні втрати формуються не в момент ураження патогеном, а значно раніше — на рівні фізіології та метаболізму рослини. Щорічні економічні втрати від абіотичних стресів обчислюються десятками мільярдів доларів, що вже сьогодні стимулює стрімке зростання ринку антистресових рішень і робить цю сферу однією з найбільш динамічних у світовій агроіндустрії. За оцінками FAO, щорічні втрати врожаю від абіотичних стресів перевищують 99 млрд доларів, і саме тому ринок антистресових рішень демонструє стрімке зростання: з 4,5 млрд доларів у 2025 році до прогнозованих 7,8 млрд доларів у 2030-му.
До базових абіотичних викликів належать посуха та нерівномірний водний режим, температурні аномалії — як перегрів, так і холодові стреси, засолення ґрунтів, порушення іонного балансу, деградація ґрунтового середовища, зсуви pH, накопичення токсичних елементів і хронічний оксидативний стрес. Принципова відмінність абіотичних факторів від біотичних полягає в тому, що вони діють постійно й кумулятивно, часто без яскраво виражених симптомів на ранніх етапах. Саме тому їхній негативний ефект довгий час залишається недооціненим, хоча він безпосередньо визначає довгостроковий потенціал урожайності.
Абіотичний стрес є системним руйнівником метаболізму рослини. Його головна небезпека полягає не лише у пригніченні росту чи тимчасовому зниженні продуктивності, а в дестабілізації базових регуляторних процесів. Під тривалим або повторюваним стресовим навантаженням порушується енергетичний баланс, знижується ефективність фотосинтезу, змінюється гормональна регуляція, накопичуються реактивні форми кисню, слабшають бар’єрні функції клітинних структур. У результаті рослина переходить із режиму розвитку в режим виживання, що обмежує реалізацію врожайного потенціалу навіть за відсутності видимих пошкоджень або симптомів.
Особливо критичним є те, що абіотичні стреси фактично «відкривають ворота» для патогенів. Ослаблений метаболізм і порушена імунна координація створюють умови, за яких біотичні агенти реалізують свою шкодочинність значно ефективніше. У багатьох виробничих ситуаціях патоген не є первинною причиною втрат, а виступає вторинним чинником, що накладається на вже ослаблений стресом організм. Саме цим пояснюється феномен, коли навіть за інтенсивного фунгіцидного або інсектицидного захисту втрати врожайності продовжують зростати — базова фізіологічна стійкість рослини залишається порушеною.
Масштаб проблеми посилюється глобальними кліматичними змінами. Зростає частота екстремальних погодних явищ, подовжуються стресові періоди, підвищується варіабельність умов навіть у межах одного сезону. За таких обставин рослина часто не має фази повноцінного відновлення, а накопичений стрес поступово знижує її адаптивний і продуктивний потенціал. Унаслідок цього втрати врожайності набувають системного, а не випадкового характеру.
Стратегічно це означає, що агроіндустрія стикається з необхідністю переосмислення підходів до управління продуктивністю. Домінування абіотичних факторів у структурі втрат свідчить про те, що фокус агротехнологій має зміщуватися від боротьби з наслідками до цілеспрямованого управління стресостійкістю. Антистресові рішення повинні працювати з базовим метаболізмом, енергетикою та імунною регуляцією рослини, а без системного підходу до абіотичного стресу контроль біотичних ризиків залишається принципово неповним. У XXI столітті саме абіотичні стреси стають центральним викликом, який визначає ефективність і конкурентоспроможність усього агровиробництва.
Чому традиційні рішення вже не працюють.
Протягом десятиліть агровиробництво розвивалося за логікою реактивного контролю: з’являється проблема — застосовується інструмент для її усунення. Фунгіциди створювалися проти грибів, інсектициди — проти шкідників, регулятори росту — для корекції морфогенезу, а системи живлення — для поповнення дефіцитів елементів. Такий підхід був ефективним у відносно стабільних кліматичних умовах минулого, однак він не враховує ключову реальність XXI століття — домінування хронічного абіотичного стресу як базового фону розвитку рослин.
Фунгіциди залишаються важливим елементом захисту, проте їхня дія принципово обмежена. Вони спрямовані виключно на патоген і не впливають на стан самої рослини. Фунгіцид не відновлює клітинний та енергетичний метаболізм, не активує системну резистентність, не підвищує толерантність до посухи, перегріву чи оксидативного навантаження і не залучає ключові регуляторні осі, такі як ABA–TOR–ROS. У реальних польових умовах препарат починає діяти вже після того, як рослина втратила частину продуктивного потенціалу через стрес. Абіотичне навантаження порушує метаболічну рівновагу, послаблює бар’єрні функції клітин і фактично «відкриває ворота» для патогенів. Фунгіцид у цій схемі є запізнілою реакцією, яка стримує розвиток хвороби, але не повертає рослину в оптимальний фізіологічний стан і не запобігає повторному ураженню. Аналогічно інсектициди впливають на шкідника, але не підсилюють адаптивні можливості рослини.
Спробою компенсувати ці обмеження стало широке впровадження біостимуляторів і антистресантів. Проте саме в цьому сегменті проявляється одна з ключових системних проблем сучасного ринку. Переважна більшість таких продуктів створюється емпірично, без чіткої молекулярної теорії дії. У більшості випадків відсутній опис того, які саме сигнальні та гормональні шляхи залучаються, як продукт впливає на баланс між ростом і стресом, чи змінює він регуляцію ABA, SA, JA, TOR або ROS-системи. Також практично відсутні комплексні дослідження, побудовані за логікою «ген — метаболізм — адаптація — урожай». У результаті ефективність біостимуляторів часто виявляється нестабільною, сильно залежить від погодних умов, фази розвитку культури та загального стресового фону. Один і той самий продукт може демонструвати позитивний ефект в один сезон і не давати відчутного результату в інший, що формує недовіру з боку виробників і ускладнює масштабування таких рішень.
У більш широкому контексті ці проблеми є наслідком того, що більшість агротехнологій досі спирається на концепцію, сформовану ще у 1990-х роках. Основу технологічних схем складають NPK-живлення, фунгіцидний та інсектицидний захист і регулятори росту. Цей набір інструментів розроблявся для умов відносно передбачуваного клімату, чіткої сезонності та обмеженої кількості стресових факторів. Сучасне ж рослинництво функціонує в умовах кліматичної нестабільності, дефіциту води, деградації ґрунтів і поєднання абіотичних та біотичних загроз упродовж усього вегетаційного періоду. Старі підходи не здатні управляти такою складною системою, оскільки вони не працюють із системною регуляцією імунітету та не враховують метаболічну пластичність рослини.
Таким чином, ключова проблема традиційних рішень полягає не в їхній «неефективності», а в обмеженості парадигми, у межах якої вони були створені. Контроль окремих факторів без управління цілісною системою адаптації рослини більше не відповідає реаліям сучасного агровиробництва. Саме це зумовлює потребу у переході до нової логіки — від реактивного захисту до проактивного управління імунітетом і стресостійкістю, що і формує основу антистресових технологій нового покоління.
Теорія рослинного імунітету: незавершеність і системний вакуум ринку.
Попри тисячі наукових публікацій і десятки років інтенсивних досліджень, рослинний імунітет досі не має завершеної, узагальненої теорії, яка пояснювала б його як цілісну керовану систему. Сучасні знання фрагментовані й описують окремі компоненти — рецептори, гормони, сигнальні каскади або реакції на стрес, але не інтегрують їх у єдину логічну модель адаптації рослини до мінливого середовища.
Наявні наукові підходи зазвичай розглядають абіотичний стрес, біотичні інфекції та регуляторні системи окремо. Імунні відповіді, пов’язані з SA, JA та етиленом, аналізуються переважно в контексті патогенів; ABA — як гормон посухи та стресу; TOR — як регулятор росту й енергетичного статусу. Водночас у реальних умовах поля всі ці системи працюють одночасно, конкуруючи за ресурси та формуючи єдину метаболічно-імунну відповідь рослини. Саме відсутність інтеграції між абіотичним стресом, біотичним тиском, гормональною регуляцією та метаболічними ресурсами (вуглецево-азотний баланс, енергетика, ROS/NO-сигнали) є ключовою науковою прогалиною.
Фактично сьогодні існує набір окремих моделей, але жодна з них не була зібрана у цілісний «метаболічно-імунний код рослини». Немає загальноприйнятого опису того, як рослина переходить між станами росту, стресу та імунної активації, де проходять межі її ресурсів і в який момент адаптація перетворюється на втрату продуктивності. Не описані чіткі зв’язки між ROS-сигналами та TOR-залежним метаболізмом, відсутні моделі порогів стійкості, після яких навіть наявність захисних препаратів уже не дає ефекту. У результаті більшість ринкових рішень не спираються на повну наукову картину дії, а використовують лише окремі фрагменти знань.

Мал. 2 Як управляти ростом і захистом рослин: новий підхід до антистресових технологій.
Цей теоретичний вакуум безпосередньо відображається на структурі сучасного аграрного ринку. Попри декларовану боротьбу зі стресом, насправді не існує механістичних препаратів, спрямованих саме на управління абіотичною адаптацією. На ринку практично відсутні продукти, здатні регулювати TOR-шлях, запускати коректний ABA-праймінг без пригнічення росту, контролювати ROS-імунітет на рівні клітинних органел або працювати з іонним гомеостазом через Na⁺/H⁺ транспортери при засоленні ґрунтів. Так само не представлені технології, які системно адаптують рослину до нестабільних температур у критичних фазах проростання та раннього розвитку.
Біостимулятори та антистресанти у своїй більшості не мають метаболічних моделей дії, тому використовуються емпірично — «на удачу», а не за чіткою формулою, що враховує стан рослини, фазу розвитку та тип стресу. Це пояснює нестабільність результатів і відсутність відтворюваності ефекту між різними сезонами й регіонами.
Відсутність інтегрованої теорії типу «енергія → сигнали → імунітет → урожай» є фундаментальним обмеженням сучасної індустрії. Саме тому підходи, які намагаються поєднати ABA-, TOR-, SA- та JA-регуляцію в єдину модель керування імунітетом, фактично перебувають на передньому краї сучасної науки й значно випереджають більшість комерційних рішень. Аграрний ринок об’єктивно чекає на системну теорію, яка дозволить перейти від фрагментарних рішень до керованої адаптації рослин, проте такої теорії на глобальному рівні досі не сформовано.
Інтегративну модель імунітету рослин проти патогенів і стресів.
Аналіз сучасних даних про молекулярні механізми формування захисних реакцій у рослин дозволив українським ученим (виконана командою науковців у складі С. Хаблака, Л. Бондаревої, М. Долі, В. Шпичака, Т. Лихолат, Т. Скляр та Ю. Лихолат) запропонувати інтегративну модель рослинного імунітету проти патогенів і стресів — «SA–JA/ET–ABA–TOR». У цій моделі імунітет рослини вперше описується не як сукупність окремих реакцій, а як цілісна динамічна сигнально-метаболічна та енергетична захисна система, керована чотирма ключовими регуляторними домінантами: саліциловою (SA), жасмоново-етиленовою (JA/ET), абсцизовою (ABA) та ростовою (TOR).
Кожна з цих домінант відповідає за певний тип загроз і стресів, а їх взаємодія визначає, як саме рослина балансує між ростом, захистом і виживанням. SA-домінанта забезпечує резистентність до біотрофних патогенів і вірусів, JA/ET — протидію некротрофам, комахам і механічним ушкодженням, ABA відповідає за адаптацію до абіотичних стресів, тоді як TOR регулює фазу росту, відновлення та продуктивності після стресових подій. Принципово новим у цій роботі стало те, що науковці вперше описали чітку ієрархію переходів між цими станами та фактори, які ними керують: енергетичний статус клітини, залучення автофагії, співвідношення вуглецю й азоту, ROS/NO-сигналізацію, мобільні РНК, а також метаболічні сигнали мікробіому.
На основі цієї інтегративної моделі імунітету рослин сформульовано практичну концепцію «Імунно-метаболічного управління культурою» — нову філософію агрономічних рішень, здатну принципово змінити підходи до захисту та живлення рослин. Це не окрема технологія і не набір рекомендацій, а цілісна модель поведінки рослини, яка пояснює, у які моменти вона перебуває в режимі росту, коли переходить до захисту, коли змушена виживати та які саме сигнали запускають ці переходи.
У центрі цієї концепції — уявлення про рослину як динамічну систему з чотирма регуляторними домінантами (SA, JA/ET, ABA і TOR), від стану яких безпосередньо залежить результат будь-якої агротехнології. Замість хаотичних або інтуїтивних обробок пропонується свідоме управління рослиною через контроль і переключення між її біологічними режимами.
Практичне застосування цієї логіки ґрунтується на чіткому розумінні функцій кожної домінанти. SA-домінанта формує системну резистентність проти біотрофів і вірусів, активує PR-гени та гіперчутливу реакцію. JA/ET-домінанта відповідає за захист від некротрофів, комах і травматичних впливів. ABA-домінанта є ключовим режимом виживання за абіотичного стресу, активуючи SnRK2-залежні каскади, антиоксидантні системи, автофагію та механізми епігенетичної пам’яті. TOR-домінанта, у свою чергу, забезпечує відновлення росту, біосинтез, роботу меристем і ефективний ресурсний метаболізм. Відповідно, агрономічне рішення має ґрунтуватися не на календарі чи стандартній схемі, а на реальному фізіологічному стані культури.
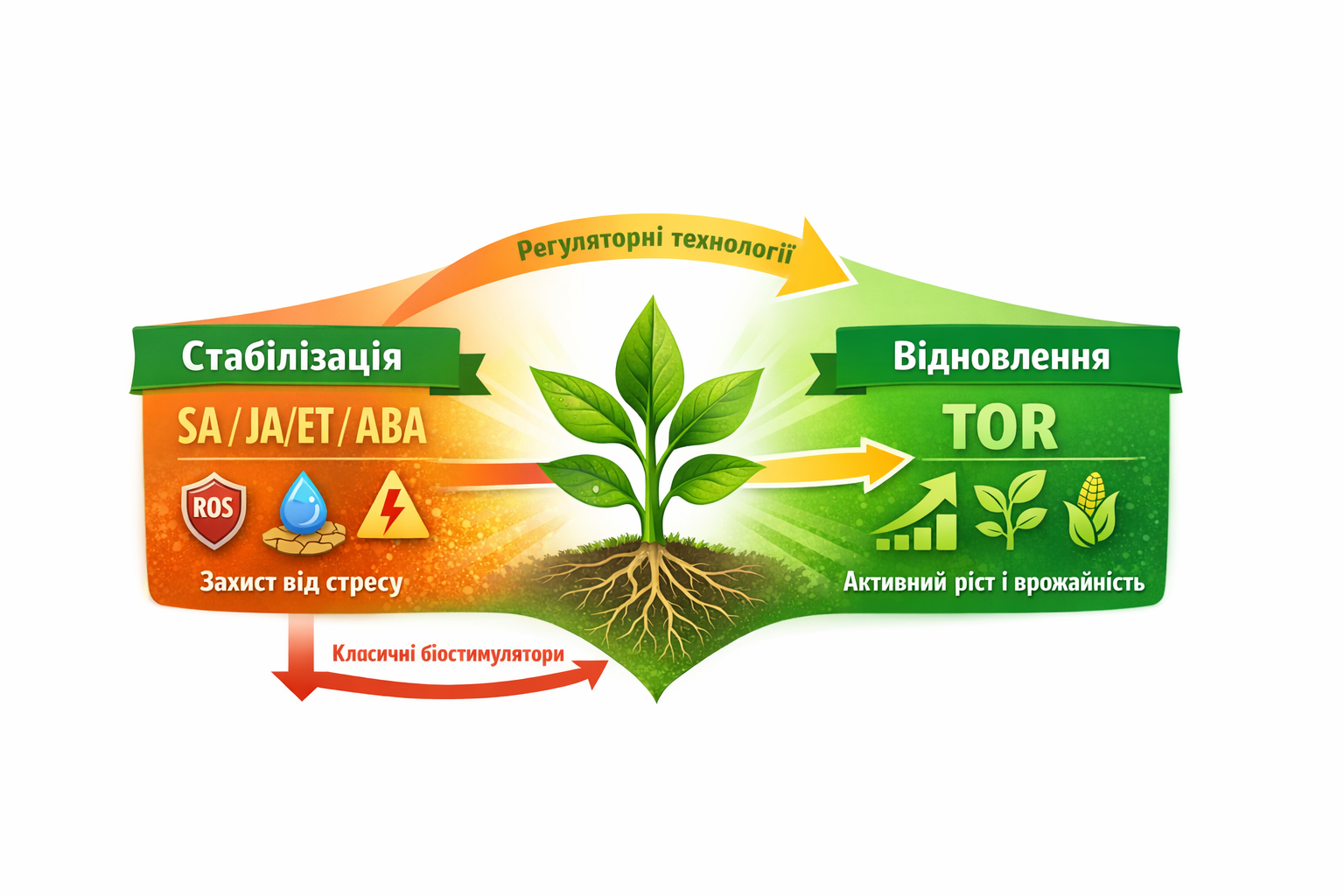
Важливим практичним наслідком цієї моделі стало формування двофазного підходу до технологій — «стабілізація → відновлення». Перша фаза передбачає ABA/SA/JA-домінантну стабілізацію, спрямовану на допомогу рослині у переживанні стресу: відновлення балансу ROS/NO, збереження тургору, зниження токсичності іонів Na⁺, Al³⁺, аміаку та важких металів, а також активацію системних сигнальних каскадів. Саме ця фаза запобігає «системному обвалу» метаболізму і дозволяє рослині зберегти потенціал урожайності.
Друга фаза — TOR-домінантне відновлення — активується після зняття або мінімізації стресу. TOR виступає центральним «комутатором» продуктивності, запускаючи біосинтетичні процеси, ріст кореня й листка, роботу меристем, відновлення білкового синтезу та подолання так званого «стресового гальма». Саме на цьому етапі формується реальне нарощування врожаю, а не лише виживання культури. Такий двофазний підхід є першою технологічною логікою, повністю узгодженою з біологією рослини, а не з інерцією традиційної агрономії.

Мал. 3 Регуляторні технології нового покоління керують стресовими та ростовими режимами рослини для максимального врожаю.
Окремим і принципово новим елементом концепції стала інтеграція мікробіому як повноцінної частини імунної системи рослини. PGPR розглядаються як активатори ABA-праймінгу та SnRK1, арбускулярні мікоризні гриби — як регулятори TOR і ресурсного балансу, ендофіти — як модератори ROS/NO-сигналів і вторинного метаболізму, а ризосферні бактерії — як стабілізатори кореневої системи й росту. Таким чином, мікробіом перестає бути «додатком» у вигляді біопрепарату і постає як функціональний орган рослини, відповідальний за її стійкість.
У підсумку концепція «Імунно-метаболічного управління культурою» є однією з наймасштабніших змін парадигми в агрономії за останні десятиліття. Вона поєднує фундаментальну біологію, епігенетику, гормональну регуляцію, ROS/NO-сигналізацію та мікробіом у єдину технологічну модель. Головне ж — вона дає агровиробникам інструмент не просто реагувати на стреси, а керувати рослиною як складною біологічною системою, здатною адаптуватися, зберігати пам’ять і свідомо перемикатися між режимами.
Розробка має потенціал стати фундаментом для нового покоління агротехнологій, здатних відповідати кліматичним викликам майбутнього. Це не лише теоретична конструкція, а практична основа для створення стресостійких сортів, точних антистресових препаратів і сучасних моделей управління врожайністю. За оцінками експертів, ця теорія може закласти підґрунтя для формування нової української школи рослинного імунітету та суттєво посилити позиції вітчизняної аграрної науки на світовій арені.
Гормональні режими SA–JA/ET–ABA–TOR у рослин: антагонізми, мультистани та помилки агротехнологій.
Сучасні рослини функціонують у складному середовищі, де одночасно діють біотичні та абіотичні стреси. Захисні та ростові реакції координуються чотирма ключовими регуляторними режимами — SA (саліцилова кислота), JA/ET (жасмонова кислота й етилен), ABA (абсцизова кислота) та TOR (Target of Rapamycin). Ці системи формують інтегровану мережу сигналізації, у якій кожен режим може повністю або частково пригнічувати інші, створюючи конфлікт між ростом і захистом.
Неправильне втручання в цей баланс через агротехнології (антистресанти, біостимулятори, фосфіти, морські водорості, амінокислоти, NPK, гормоноактивні препарати) часто зумовлює десинхронізацію імунних станів, що призводить до суттєвих втрат врожаю — від 10 до 45%.
Центральною особливістю інтегративної моделі рослинного імунітету є те, що регуляторні системи SA, JA/ET, ABA і TOR не працюють паралельно або адитивно. Навпаки, між ними існує жорстка мережа антагонізмів, яка дозволяє рослині швидко й однозначно переходити між різними біологічними режимами — ростом, імунним захистом або виживанням. Саме ці антагонізми формують логіку прийняття «рішень» рослиною під час стресу чи інфекції.
Саліцилова кислота активується переважно у відповідь на біотрофні патогени (бактерії, біотрофні гриби) та вірусні інфекції. Її включення переводить рослину у режим так званої оборони першого типу, що характеризується локалізацією інфекції, запуском гіперчутливої реакції, індукцією PR-білків і зміцненням клітинних стінок. Для ефективності такої відповіді необхідне жорстке перепрограмування клітини, і тому SA активно пригнічує інші регуляторні шляхи. Зокрема, вона блокує JA/ET-сигналізацію, оскільки жасмоново-етиленові програми сприяють некротизації тканин, ROS-вибуху та ослабленню клітинних стінок — процесам, які є вигідними для некротрофів, але небезпечними у випадку біотрофної інфекції. Паралельно SA пригнічує TOR-сигналізацію, фактично зупиняючи ріст і переводячи енергетичні та метаболічні ресурси з біосинтезу на імунний захист. У SA-домінантному стані рослина свідомо жертвує ростом заради локалізації загрози.
Жасмоново-етиленова регуляція, навпаки, активується при некротрофних інфекціях (некротрофні гриби), пошкодженнях тканин, жуванні комахами та механічному стресі (град). Цей шлях запускає інший тип імунної відповіді, орієнтований на агресивний захист, що включає програмовану загибель клітин, посилення дихання та інтенсивну ROS-сигналізацію. У такому контексті JA/ET пригнічує SA-систему, оскільки SA спрямована на збереження клітин і локалізацію інфекції, що суперечить логіці боротьби з некротрофами. Блокування SA відбувається через репресію NPR1/EDS1-залежних каскадів і зниження накопичення саліцилової кислоти. Паралельно JA/ET пригнічує ABA, оскільки абсцизова кислота переводить рослину у водоощадний і метаболічно пригальмований режим, тоді як ефективний захист від некротрофів вимагає високих енергетичних витрат і активного метаболізму.
Абсцизова кислота є ключовим регулятором відповіді на абіотичні стреси, такі як посуха, холод або теплове навантаження. Активація ABA переводить рослину у режим виживання, у якому пріоритетом стає збереження води, енергії та структурної цілісності клітин. У цьому стані ABA системно пригнічує як SA-, так і JA/ET-залежні програми, оскільки імунні реакції є надзвичайно енергоємними та супроводжуються підвищеним утворенням ROS, що за умов стресу може призвести до незворотних пошкоджень. На молекулярному рівні ABA знижує активність EDS1/NPR1-комплексу, пригнічує синтез PR-білків і обмежує JA/ET-індуковані каскади. Одночасно ABA через активацію SnRK2-кіназ ефективно вимикає TOR-сигналізацію, зупиняючи ріст і біосинтез. Таким чином, ABA-домінанта є «жорстким перемикачем» у режим економії ресурсів і виживання.
TOR, у свою чергу, виступає центральним регулятором росту й продуктивності. Його активація сигналізує про сприятливі умови, достатній рівень енергії та поживних речовин. У TOR-домінантному стані рослина стимулює синтез білків, амінокислот і нуклеїнових кислот, активує меристеми та підтримує високий метаболічний потік. Для збереження цього режиму TOR активно пригнічує стресові й імунні сигнали, зокрема ABA та SA. Це пригнічення запобігає переходу рослини у режим економії або імунної мобілізації, які неминуче обмежують ріст. TOR також знижує рівень автофагії та ROS-сигналізації, «стираючи» стресові сигнали і закріплюючи стан активного розвитку.
У сукупності ці антагоністичні взаємодії формують чітку ієрархію регуляторних станів і пояснюють, чому рослина не може одночасно максимально рости, активно захищатися і переживати стрес. Кожна домінанта не лише активує власні програми, а й цілеспрямовано вимикає альтернативні шляхи. Саме ця логіка лежить в основі інтегративної моделі «SA–JA/ET–ABA–TOR» і дозволяє перейти від фрагментарного опису реакцій до системного розуміння того, як рослина приймає біологічні рішення в умовах мінливого середовища.
Таблиця 1
Антагонізми та інтеграція регуляторних шляхів SA, JA/ET, ABA і TOR у рослині
| Домінуючий регулятор | Основний тригер | Що активує | Що пригнічує | Функціональний режим рослини | Біологічна логіка |
| SA (саліцилова кислота) | Біотрофні патогени | PR-білки, HR, зміцнення клітинної стінки | JA/ET, TOR | Локальний імунітет проти біотрофів | Запобігання некротизації та перенаправлення ресурсів з росту в захист |
| JA/ET (жасмонова + етилен) | Некротрофи, пошкодження, комахи | PCD, ROS, некротичний захист | SA, ABA | Агресивний імунітет проти некротрофів | Некротрофи виграють від живих клітин → потрібна контрольована загибель |
| ABA (абсцизова кислота) | Посуха, холод, перегрів | Контроль продихів, SnRK2 | SA, JA/ET, TOR | Режим виживання та енергозбереження | Обмеження дорогих імунних програм і росту під стресом |
| TOR (Target of Rapamycin) | Достаток ресурсів, енергії | Біосинтез, ріст, поділ клітин | ABA, SA | Ростовий режим | Стресові та імунні програми блокують ріст — TOR їх пригнічує |
Таблиця 2
Регулятор → Антагонізм → Метаболічний наслідок
| Регулятор | Антагонізм | Прямий наслідок | Метаболічний ефект | Ризик при неправильній активації |
| SA | ⟂ JA/ET | Блок некротизації | Збереження клітин | Вразливість до некротрофів |
| SA | ⟂ TOR | Зупинка росту | Перерозподіл ресурсів | Втрата врожайності |
| JA/ET | ⟂ SA | Блок PR-білків | Посилення ROS | Надлишковий некроз |
| JA/ET | ⟂ ABA | Відкриті продихи | Високий метаболізм | Втрата води |
| ABA | ⟂ TOR | Стоп росту | Енергозбереження | Затримка розвитку |
| TOR | ⟂ ABA | Активний ріст | Високий C/N обмін | Зниження стійкості |
| TOR | ⟂ SA | Менше імунітету | Пріоритет біосинтезу | Вразливість до інфекцій |
Таблиця нижче відображає ключові біологічні режими рослини та їх взаємодію між ростом, імунітетом і стресовими сигналами. Це допомагає зрозуміти, чому універсальних «антистресантів» не існує та чому кожне втручання в рослину — це не просто стимуляція всього, а зміщення балансу між ростом і захистом. Вона також показує, де сучасний ринок є «сліпим», потребує науково обґрунтованих рішень і закладає основу для створення механістичних продуктів нового покоління.
Таблиця 3
Регуляторні режими як «стани рослини»
| Стан рослини | Домінуючі сигнали | TOR | Імунітет | Ріст | Типовий ризик |
| Ростовий | TOR ↑ | Активний | Мінімальний | Максимальний | Інфекції |
| Біотрофний імунітет | SA ↑ | Пригнічений | Високий (локальний) | Низький | Втрата біомаси |
| Некротрофний імунітет | JA/ET ↑ | Частково активний | Агресивний | Середній | Некроз |
| Абіотичний стрес | ABA ↑ | Вимкнений | Пригнічений | Мінімальний | Втрата потенціалу |
| Перехідний / адаптивний | ABA + TOR (контрольовано) | Баланс | Праймінг | Контрольований | — |
Природа конфлікту: захист проти росту.
Рослина не здатна одночасно підтримувати максимальний ріст і повноцінний захист. Це не агрономічна умовність, а фундаментальний біологічний принцип, закладений у саму архітектуру регуляторних систем. На молекулярному рівні ріст і захист керуються антагоністичними сигнальними режимами, які працюють як взаємовиключний перемикач станів.
Режим росту контролюється комплексом TOR (Target of Rapamycin). Його активація означає пріоритет біосинтезу: інтенсивний синтез амінокислот, білків і нуклеїнових кислот, активний поділ клітин, високий рівень фотосинтетичного потоку та максимальне використання азоту й енергії. У цьому стані рослина оптимізована на накопичення біомаси й формування врожайності, але імунні та стресові програми свідомо пригнічені як метаболічно надто дорогі.
На протилежному боці знаходяться режими захисту, які запускаються гормональними сигналами SA, JA/ET та ABA. Незважаючи на різну специфіку тригерів — біотрофні патогени, некротрофи, механічні ушкодження або абіотичні стреси — всі ці сигнали переводять рослину у стан стресової адаптації. У цьому режимі ресурси перенаправляються з росту в оборону: активуються імунні каскади, посилюється клітинна стінка, індукується контрольований ROS-сигналінг, обмежується поділ клітин і пригнічується TOR-залежний біосинтез.
Фізіологічно це супроводжується уповільненням споживання азоту, перебудовою фотосинтезу з продуктивного на підтримуючий, зниженням темпів росту та, у випадку ABA, жорстким режимом енергозбереження. Така стратегія критично важлива для виживання, але несумісна з високою продуктивністю.
Саме тут виникає ключовий конфлікт: неправильний або несвоєчасний вибір регуляторного режиму має високу ціну. Передчасна або надмірна активація стресових шляхів призводить до втрати потенційної врожайності навіть за відсутності реальної загрози. Водночас збереження ростового режиму TOR в умовах патогенного чи абіотичного тиску робить рослину вразливою до інфекцій, пошкоджень і незворотних втрат.

Мал. 4 Системний контроль стресостійкості культур через імунно-метаболічне управління.
Цей конфлікт лежить в основі відмінності між класичними біостимуляторами та сучасними регуляторними технологіями. Перші працюють за принципом «підштовхнути рослину до росту», незалежно від її стану. В умовах стресу це часто призводить до неефективності: активний TOR у момент, коли рослина повинна стабілізуватися, несе ризик енергетичного дефіциту, ROS-дисбалансу і ослаблення імунітету.
Регуляторні технології нового покоління діють за зовсім іншою логікою. Вони керують перемиканням режимів рослини, починаючи зі стабілізації стресових домінант (SA, JA/ET, ABA), відновлюючи метаболічну і гормональну рівновагу, і лише потім активують TOR для відновлення росту та продуктивності. Такий підхід відповідає природній фізіології рослини і дозволяє одночасно мінімізувати втрати врожайності та посилити стресостійкість культури.
Класичні біостимулятори сформувалися в парадигмі, де рослина розглядається як пасивний об’єкт, якому достатньо «додати ресурс» для посилення росту. У такій логіці амінокислоти, гумінові речовини або мікроелементи мають автоматично трансформуватися в біомасу та врожайність. Проте сучасна фізіологія рослин показує, що за умов стресу ця логіка не працює: рослина цілеспрямовано блокує ростові програми через пригнічення TOR-сигналізації та переключається в режими виживання, керовані SA, JA/ET або ABA.
Регуляторні технології виходять з протилежної логіки: рослина — це активна саморегульована система з чіткими біологічними режимами. Їхнє завдання — не стимулювати ріст за будь-яку ціну, а керувати переходами між режимами «стрес → стабілізація → відновлення». Саме тому в таких технологіях первинною є фаза стабілізації (ABA/SA/JA), і лише після відновлення метаболічної рівноваги запускається TOR-залежне нарощування продуктивності.
Класичні біостимулятори не враховують антагонізм між ростовою TOR-системою та стресовими сигнальними домінантами SA, JA/ET і ABA. За наявності абіотичного або біотичного стресу активація TOR є біологічно забороненою, оскільки призводить до енергетичного дефіциту, ROS-дисбалансу та зниження стійкості. У таких умовах будь-яка спроба стимулювати ріст шляхом зовнішнього внесення ресурсів має низьку ефективність або навіть погіршує стан культури.
Регуляторні технології діють на рівні сигнальних каскадів: вони коригують гормональну і метаболічну домінанту, стабілізують ROS/NO-сигнали, оптимізують енергетичний баланс і лише після цього дозволяють активацію TOR. Таким чином, продуктивність не нав’язується рослині, а стає природним наслідком правильного фізіологічного стану.
Класичні біостимулятори працюють так, ніби людину з температурою, зневодненням і запаленням намагаються змусити бігти марафон, просто давши їй більше їжі. Регуляторні технології спочатку «лікують» — знімають стрес, стабілізують систему, відновлюють баланс, і лише потім повертають організм до повноцінної роботи та високої продуктивності.
Таким чином, конфлікт «ріст проти захисту» є не проблемою окремих гормонів чи факторів, а системною властивістю рослинного метаболізму. Ефективне управління продуктивністю можливе лише через точне керування переходами між цими станами, а не через спроби одночасно стимулювати взаємовиключні біологічні програми.
Ключова відмінність між класичними біостимуляторами та регуляторними технологіями полягає у філософії впливу на рослину. Перші намагаються стимулювати ріст незалежно від фізіологічного стану культури, тоді як другі керують біологічними режимами рослини. Регуляторний підхід передбачає первинну стабілізацію стресових домінант (SA, JA/ET, ABA) і лише після цього — контрольоване відновлення росту через TOR. Саме така послідовність відповідає природній логіці функціонування рослин і забезпечує стабільну реалізацію врожайного потенціалу.
Мал. 5 Двофазна модель регуляції рослини: стабілізація через SA/JA/ET/ABA → відновлення росту через TOR. Порівняння класичних біостимуляторів і сучасних регуляторних технологій.
Таблиця 4
Класичні біостимулятори у порівнянні з регуляторними технологіями
| Критерій | Класичні біостимулятори | Регуляторні технології нового покоління |
| Базова логіка | «Дати рослині більше» (амінокислоти, мікроелементи, гумати) | «Перевести рослину в правильний біологічний режим» |
| Розуміння фізіології | Загальне, без урахування гормональних конфліктів | Ґрунтується на антагонізмах SA–JA/ET–ABA–TOR |
| Робота зі стресом | Маскування симптомів | Управління стресовими домінантами |
| Вплив на TOR | Часто хаотично активують TOR навіть під стресом | Чітко розмежовують: стабілізація → TOR-відновлення |
| Ріст під стресом | Стимулюється штучно | Тимчасово блокується заради виживання |
| Імунна відповідь | Неспецифічна або відсутня | Спрямована (SA, JA/ET або ABA залежно від загрози) |
| Урахування типу патогена | Не враховується | Біотрофи / некротрофи / абіотика — різні рішення |
| Робота з ROS/NO | Побічний ефект | Центральний механізм перемикання режимів |
| Енергетичний баланс | Ігнорується | Ключовий фактор прийняття рішень |
| Автофагія та відновлення | Не враховуються | Свідомо керовані фази |
| Роль мікробіому | “Корисне доповнення” | Повноцінний елемент імунної системи |
| Час застосування | За календарем або фазою | За фізіологічним станом культури |
| Результат | Нестабільний, залежний від умов | Передбачуваний і відтворюваний |
| Ризики | Високі при стресах | Мінімізовані за рахунок біологічної логіки |
| Філософія | Реактивна | Проактивна, системна |
| Покоління підходу | Агрохімічна логіка XX століття | Імунно-метаболічна логіка XXI століття |
Що відбувається, коли аграрій застосовує «не ті» речовини в «не той» імунний стан рослини.
Ключова проблема сучасних агротехнологій полягає в тому, що більшість рішень застосовуються за календарем, а не за фізіологічним станом рослини. При цьому рослина в кожен момент часу перебуває лише в одному домінантному регуляторному режимі — ростовому або стресово-імунному. Будь-яке втручання, яке суперечить цьому режиму, не просто не допомагає, а часто різко погіршує ситуацію.
Рослинний імунітет працює за принципом виключного перемикання: TOR відповідає за ріст і продуктивність, тоді як SA, JA/ET та ABA активують різні форми захисту і стрес-адаптації. Помилка у виборі режиму означає або штучне вимикання імунітету, або безпідставне гальмування росту. Саме на цьому рівні виникають ключові агротехнологічні втрати.
Однією з найпоширеніших помилок є активація TOR під час біотрофної інфекції, коли рослина природно переходить у SA-режим. Типова ситуація — після дощу, на початку розвитку іржі або борошнистої роси, по листу вносять амінокислоти, гумати та NPK. У результаті TOR починає домінувати над SA-сигналами, і рослина фактично вимикає власну оборону. Біотроф отримує ідеальні умови для розмноження, а втрати врожайності можуть сягати 20–40%. Формально «живлення» присутнє, але імунітет уже зламаний.
Не менш небезпечною є активація JA/ET під час посухи, коли рослина перебуває в ABA-режимі. ABA намагається зберегти воду, зменшити транспірацію та стабілізувати клітинні мембрани. Натомість JA/ET різко підвищують рівень ROS, активують катаболічні процеси та прискорюють старіння. Коли в умовах водного дефіциту застосовують водорості, цитокініни або агресивні стимулятори, виникає різка десинхронізація регуляції: ABA блокується, рослина втрачає тургор, з’являються опіки листя та передчасне старіння. Типові втрати в таких сценаріях становлять 10–30%.
Третя системна помилка — застосування SA-активаторів проти некротрофів, коли ключовим має бути JA/ET-режим. Наприклад, використання фосфітів або саліцилатних схем на соняшнику проти Alternaria чи Sclerotinia. SA блокує JA/ET-сигнали, а саме вони відповідають за ефективний захист від некротрофів. У результаті некротроф отримує перевагу, локальні некрози посилюються, а ураження може зростати в 2–4 рази. Залежно від культури, втрати врожайності сягають 10–35%.
Зворотна, але не менш шкідлива ситуація — активація ABA у фазі активного росту, коли рослина перебуває в TOR-режимі. Завчасне застосування антистресантів, водоростевих екстрактів або гуматів без реального стресу призводить до пригнічення TOR. Фотосинтез знижується, коренева система росте повільніше, ефективність використання азоту падає. У підсумку рослина втрачає 5–15% продуктивного потенціалу без жодної реальної потреби.
Найбільш руйнівною практикою є змішування «всього з усім» — так звані агрономічні коктейлі. Амінокислоти (TOR), морські водорості (частково ABA), фосфіти (SA), піретроїди (інсектицид-індукований JA/ET-стрес) часто поєднуються в одному баку без будь-якої логіки. У такій системі сигнали не підсилюють одне одного, а взаємно блокуються. Рослина входить у стан метаболічного хаосу: і ріст, і імунітет пригнічені, ресурс витрачається неефективно, а втрати врожайності коливаються від 10 до 40%.
Таким чином, проблема сучасних технологій полягає не у відсутності препаратів, а у відсутності розуміння регуляторних режимів рослини. Поки агротехнології не перейдуть від календарної логіки до керування станами — «ріст → стрес → імунітет → відновлення» — навіть найкращі продукти працюватимуть нестабільно або проти самої рослини.
Як це проявляється у сучасних технологіях вирощування.
У більшості сучасних систем вирощування ключова помилка полягає не у виборі окремих препаратів, а у відсутності діагностики імунного та метаболічного стану рослини в момент втручання. Технологічні рішення приймаються “за календарем”, за фазою розвитку або за стандартною схемою, тоді як сама рослина вже перебуває у певному регуляторному режимі — ростовому, імунному або стресовому. Ігнорування цього факту створює глибокий дисонанс між внутрішньою сигналізацією рослини та зовнішнім агрономічним впливом.
На практиці це проявляється у масовому застосуванні антистресантів у періоди, коли реального абіотичного стресу немає, стимуляторів росту під час початкових стадій інфекції, фосфітів проти некротрофних патогенів, цитокінінів у фазі водного дефіциту або амінокислот у період активного патогенного тиску. Кожне з таких рішень саме по собі може виглядати логічним, однак у сукупності вони порушують узгодженість між SA-, JA/ET-, ABA- та TOR-регуляторними режимами, створюючи конфлікт сигналів усередині клітини.
Наслідком цього гормонального і метаболічного дисбалансу стає не миттєва катастрофа, а системне “просідання” фізіології культури. Зростає частка латентних інфекцій, які не проявляються одразу, але поступово знижують продуктивність. Посилюється передчасне старіння листкового апарату, зменшується коефіцієнт використання азоту, падає індекс листкової поверхні, слабшає розвиток кореневої системи, а формування генеративних органів відбувається в умовах обмеженого ресурсу. У результаті рослина формально “захищена” або “стимульована”, але функціонально — дезорганізована.
Саме тому у багатьох господарствах фіксуються стабільні втрати врожайності на рівні 15–45%, а в умовах комбінованого стресу — ще вищі. Ці втрати не є наслідком одного неправильного препарату, а результатом системної помилки: невміння працювати з рослиною як із динамічною біологічною системою, де кожне втручання має відповідати її поточному імунно-метаболічному стану.
Мультистани: коли одночасно активні кілька режимів.
У реальних польових умовах рослина майже ніколи не перебуває у «чистому» ростовому або імунному режимі. Набагато частіше вона змушена одночасно реагувати на кілька факторів — наприклад, на поєднання біотрофної та некротрофної інфекції, посухи з патогенним тиском або високих температур із механічними пошкодженнями. Такі ситуації формують так званий мультистан — компромісний метаболічний режим, у якому активуються одразу кілька сигнальних систем. Рослина знаходиться у компромісному стані, де жодна програма повністю не домінує. Це метаболічно затратний стан, і рослина не може ефективно рости і одночасно боротися з усіма стресами. Дана ситуація реалістична в полі, особливо коли рослина піддається комбінованому стресу: одночасно присутні біотрофи та некротрофи, водний стрес і механічне пошкодження, сильне сонце та патогенна атака.
Таблиця 5
Як проявляються мультистани на рівні гормональних систем
| Гормон / сигнал | Можливий стан у мультистані | Наслідки для рослини |
| SA | частково активний | оборона проти біотрофів активна, але пригнічена JA/ET |
| JA/ET | частково активний | захист проти некротрофів частково працює, але пригнічує SA |
| ABA | високий через абіотичний стрес | пригнічує SA та JA/ET, гальмує TOR |
| TOR | частково активний, якщо умови росту сприятливі | стимулює ріст, але конфліктує з ABA і SA |
Мультистани — це природний стан рослини при комплексних стресах. Головне завдання агронома — визначити пріоритети та баланс, застосовувати препарати помірно та поетапно, щоб уникнути конфлікту між SA, JA/ET, ABA та TOR.
У мультистані саліцилатна (SA) та жасмонат-етиленова (JA/ET) сигналізація працюють частково й нерівномірно, абсцизова кислота (ABA) зазвичай перебуває на високому рівні, намагаючись зберегти воду й стабільність клітин, а TOR-сигналінг не вимикається повністю, але переходить у обмежений, фрагментований режим. У результаті рослина опиняється в енергетичному дефіциті: ресурсів не вистачає ні для повноцінного захисту, ні для активного росту. Імунна відповідь залишається неповною, ріст сповільнюється, а ризик прискореного старіння різко зростає.
Саме мультистани є найбільш уразливими до агротехнологічних помилок, оскільки будь-яке різке втручання — надмірна стимуляція TOR, агресивна активація SA або JA/ET чи передчасне підсилення ABA — може зруйнувати крихкий баланс і перевести рослину в стан глибокого фізіологічного виснаження.
У таких умовах роль агронома змінюється: він уже не «стимулює» культуру, а керує пріоритетами. Першим принципом стає пріоритетність стресів — спочатку нейтралізується найбільш загрозливий фактор. Якщо домінує патогенний тиск, фокус робиться на відповідному імунному контурі (SA для біотрофів або JA/ET для некротрофів); якщо ключовим є абіотичний стрес, першочергово стабілізується ABA-залежна регуляція. Лише після цього можливе обережне повернення до відновлення ростових процесів через контрольовану активацію TOR. Спочатку усувається найнебезпечніший стрес: патогенний → SA або JA/ET, абіотичний → ABA, після стабілізації → TOR.
Другим принципом є використання м’яких мультисигнальних рішень — низькоконцентрованих комплексів, які підтримують баланс між режимами, а не різко зсувають рослину в один із них. Такі підходи знижують ризик гормональних «зламів» і дозволяють рослині самостійно інтегрувати сигнали відповідно до поточного стану.
Третім критично важливим елементом стає поетапність внесення. У мультистанах ефективні не «одним махом» рішення, а послідовна логіка: спочатку стабілізація (антистрес або ABA-орієнтовані впливи), далі — корекція імунної відповіді (SA або JA/ET залежно від типу загрози), і лише після цього — обережне відновлення росту через TOR-сигналінг. По днях – день 1: ABA або антистрес, день 2: SA або JA/ET, день 3: TOR (відновлення). Така тимчасова декомпозиція дозволяє рослині вийти з кризового режиму без втрати продуктивного потенціалу.
Основою ж усього цього підходу є діагностика. Без розуміння водного статусу, наявності латентної інфекції, швидкості росту та ступеня пошкоджень будь-яка технологія залишається гаданням. Мультистани вимагають не універсальних рецептів, а точного зчитування фізіологічного стану рослини й регуляторно вивірених рішень, які працюють не проти її біології, а разом із нею.
Значення правильного керування режимами SA–JA/ET–ABA–TOR.
Продуктивність рослини визначається не кількістю внесених препаратів, а тим, наскільки точно агротехнологія узгоджена з її внутрішніми регуляторними режимами. Системи SA, JA/ET, ABA та TOR не є «стимуляторами» або «антистресантами» у класичному розумінні — це базові перемикачі фізіологічних станів, які визначають, куди саме рослина спрямовує свої ресурси: на захист, виживання або ріст.
Саліцилатний режим (SA) є ключовим для захисту від біотрофних патогенів. Його активація дозволяє локалізувати інфекцію, але водночас пригнічує ростові програми та TOR-залежний метаболізм. Жасмонатно-етиленовий контур (JA/ET) відповідає за боротьбу з некротрофами та реакцію на механічні пошкодження; він більш «агресивний», супроводжується підвищенням рівня ROS і також має антагоністичні відносини з ростом. Абсцизова кислота (ABA) переводить рослину в режим виживання під абіотичним стресом — посухою, холодом, засоленням або перегрівом, зберігаючи воду й клітинну стабільність ціною тимчасової зупинки росту. TOR-сигналінг, навпаки, є центром росту та відновлення, що активує синтез білків, клітинний поділ і формування врожаю, але працює ефективно лише тоді, коли стресові сигнали знижені.
Критично важливо розуміти, що ці режими не додаються один до одного механічно. Їхнє хаотичне змішування без урахування поточного стану культури призводить до імунологічних і метаболічних помилок: блокування захисних контурів, передчасного старіння, втрати води, зниження ефективності використання азоту та енергії. У таких умовах потенціал урожаю зменшується ще до того, як починають працювати пестициди, добрива або фунгіциди.
Саме тому правильна робота з режимами SA, JA/ET, ABA та TOR є стратегічною основою сучасних технологій вирощування. Вона дозволяє не просто реагувати на проблеми, а керувати переходами між станами — від захисту й виживання до контрольованого відновлення росту. Там, де сигнали узгоджені, рослина використовує ресурси ефективно і реалізує генетично закладений потенціал. Там, де режими плутаються, втрати можуть сягати 30–45% навіть за формально «повної» системи захисту та живлення.
Наведена нижче таблиця узагальнює ключову логіку регуляції продуктивності рослин: кожен гормональний або імунний стан відповідає конкретному біологічному режиму, і втручання в цей режим повинно бути строго узгоджене з ним. Сигнальні системи SA, JA/ET, ABA та TOR не є адитивними — вони працюють антагоністично й конкурують за енергетичні та метаболічні ресурси рослини. Саме тому спроби одночасно стимулювати ріст і захист або «підстрахуватися всім одразу» призводять не до синергії, а до дезорганізації регуляції.
Таблиця наочно показує, що неправильний вибір речовин і моменту їх застосування здатен знизити врожайність на 10–45% ще до того, як почнуть діяти патогени або спрацюють пестициди. Водночас правильне узгодження препаратів із домінуючим імунним чи фізіологічним станом рослини дозволяє зберегти ресурси, підтримати адекватний захист і забезпечити контрольоване відновлення росту.
Таким чином, оптимальна агротехніка базується не на календарному графіку внесень, а на діагностиці стану рослини — фазі росту, типі стресу та характері патогенного тиску. Саме цю логіку «стан → сигнал → рішення → результат» і відображає наведена нижче таблиця.
Таблиця 6
“Правильно / неправильно” застосування стимуляторів/гормональних комплексів залежно від імунного стану рослини (SA, JA/ET, ABA, TOR):
| Імунний / гормональний стан рослини | Бажана дія / речовини | Правильне застосування | Неправильне застосування та наслідки |
| SA-домінантний (біотрофи) | Саліцилати, фосфіти, біоконтроль проти біотрофів | Підсилює PR-білки, локалізує інфекцію, економно витрачає ресурси | TOR-стимулятори, амінокислоти → пригнічують SA → біотрофи швидко розмножуються, зниження захисту |
| JA/ET-домінантний (некротрофи, пошкодження) | JA-аналоги, ET-індуктори, механічна стимуляція | Активує ROS, PCD, оборону проти некротрофів і травм | SA-стимулятори → блокують JA/ET, посилюється ураження некротрофами; ABA → пригнічує JA/ET → слабкий захист |
| ABA-домінантний (посуха, холод, стрес) | ABA-подібні антистресанти, осмотики, контроль продихів | Підвищує виживання, економить воду, зберігає клітини | TOR / JA/ET / SA-стимулятори → метаболічний дисонанс, ROS, передчасне старіння, втрати тургору |
| TOR-домінантний (активний ріст, оптимальні умови) | Амінокислоти, гумати, NPK, мікроелементи | Стимулює синтез білків, ДНК, ріст, фотосинтез | ABA → пригнічує TOR, зменшує ріст; SA → пригнічує TOR → ростові програми гальмуються |
Індустрія потребує наукових консультантів нового покоління.
Сучасний аграрний ринок увійшов у фазу, коли традиційних ролей — агронома-практика, продавця препаратів чи окремого науковця — вже недостатньо. Зростання складності стресів, мультифакторних уражень і нестабільних сезонів вимагає фахівців нового типу, здатних мислити системно й працювати на перетині кількох дисциплін одночасно.
Індустрії потрібні консультанти, які можуть інтегрувати імунологію рослин, фізіологію, біохімію та практичну агрономію в єдину логіку рішень. Такі фахівці не просто радять окремі продукти, а формують технологічні карти за фазами розвитку культури й за її біологічними станами — від стресової стабілізації до контрольованого відновлення росту. Вони здатні вибудовувати нову, науково обґрунтовану систему підбору антистресантів і стимуляторів, де кожне втручання має чітку механістичну мету.
Ключовою відмінністю консультанта нового покоління є вміння пояснювати складні молекулярні механізми простою, зрозумілою мовою — як фермеру, так і маркетингу чи управлінській команді. Його рекомендації базуються не на загальних формулюваннях і рекламних тезах, а на метаболічній моделі рослини, розумінні антагонізмів сигнальних шляхів і реальних обмежень ресурсу.
Саме таких фахівців сьогодні критично бракує ринку. І саме вони стають ключовою ланкою між фундаментальною наукою, технологіями нового покоління та практичною ефективністю агровиробництва.

Мал. 6 Нова логіка внесення антистресантів: від календаря до стану рослини.
Висновки
Висока та стабільна продуктивність рослин напряму залежить від того, наскільки грамотно керуються їхні внутрішні гормональні режими. Сучасні втрати врожайності найчастіше виникають не через нестачу препаратів чи елементів живлення, а через ігнорування ключового питання — в якому імунному та метаболічному стані перебуває рослина в момент втручання. Коли агротехнологія працює «по календарю», а не по фізіології, навіть правильні за складом продукти можуть запускати протилежні процеси й послаблювати культуру.
Розуміння антагонізмів і взаємодій між SA, JA/ET, ABA та TOR дає можливість будувати технології нового типу — точні, логічні, прогнозовані та енергоефективні. Такі підходи не стимулюють «усе одразу», а свідомо керують переходами між режимами захисту, виживання та росту, зберігаючи ресурсний баланс рослини.
Особливої ваги набуває комплексне бачення мультистанів, у яких культура одночасно зазнає кількох стресів. Саме тут відкривається потенціал для інтелектуальних схем внесення — адаптивних, поетапних і механістично обґрунтованих. Вони дозволяють мінімізувати конфлікти сигналів, зменшити приховані втрати та стабільно реалізувати урожайний потенціал навіть у нестабільних умовах вирощування.
Хаблак Сергій, д.б.н., агроном-експерт, агроаудитор
Читайте нас у Telegram









Sumitomo Chemical подала заявки на реєстрацію нового фунгіциду bifemetstrobin у Японії, США та Бразилії
13.07.2026, 11:27
ЗЗР та насіння

Італійські дослідники підтвердили ефективність дріжджового біопрепарату проти несправжньої борошнистої роси винограду
13.07.2026, 09:07
ЗЗР та насіння

Дослідники виявили перспективний біоагент проти фузаріозного в’янення бананів
12.07.2026, 23:41
ЗЗР та насіння

При купівлі вершкового масла приділяйте увагу інформації на маркуванні
12.07.2026, 10:49
Новини