
Практичне управління енергією та стресом: інструменти збереження врожаю
16.03.2026

ЧАСТИНА ІІ.
УРОЖАЙ — ЦЕ НАУКА В ДІЇ.
КОНТРОЛЮЙ СТРЕС — КОНТРОЛЮЙ ВРОЖАЙ.

Мал. 1. Прояви гербіцидного стресу на рослинах кукурудзи, спричиненого застосуванням гормональних гербіцидів із порушенням рекомендованого температурного режиму, що призводить до фізіологічної дезрегуляції, пригнічення ростових процесів та тимчасового розбалансування метаболізму.
У першій частині було сформовано теоретичну основу енергетичної моделі стресу. Показано, що стрес — це насамперед порушення енергетичного балансу клітини, пригнічення фотосинтезу та перемикання рослини з режиму росту (TOR-домінантний стан) у режим виживання (SnRK1/ABA-домінантний стан). Було обґрунтовано, що саме стан хлоропластів і рівень енергетичної забезпеченості визначають, чи зможе рослина реалізувати свій генетичний потенціал продуктивності.
Друга частина переходить від теорії до практики. Її мета — показати, як саме керувати енергією, фотосинтезом, ROS та метаболічними перемикачами в польових умовах, щоб не допустити енергетичного колапсу клітини та втрати врожайності.
Як підтримати фотосинтез — головне джерело енергії?
Підтримання ефективного фотосинтезу є одним із ключових факторів виживання та продуктивності рослин, особливо в умовах дії абіотичних і біотичних стресів. Саме фотосинтез забезпечує утворення АТФ, відновлювальних еквівалентів і вуглецевих сполук, які є основою для всіх процесів росту, розвитку і формування врожаю. Центральним елементом цього процесу є хлоропласти, які виконують не лише функцію синтезу органічних речовин, але й виступають важливими сенсорами стресу. За несприятливих умов, таких як посуха, низькі або високі температури, гербіцидне навантаження чи дефіцит елементів живлення, робота фотосинтетичного апарату порушується, що призводить до зниження синтезу енергії та накопичення реактивних форм кисню (ROS), які додатково пошкоджують клітинні структури. Тому одним із головних практичних завдань сучасної агрономії є збереження функціональної активності хлоропластів і стабільності фотосинтезу в умовах стресу.
Вирішальну роль у підтриманні стабільності фотосинтетичного апарату відіграє магній, який є центральним атомом молекули хлорофілу і безпосередньо визначає здатність рослини поглинати світлову енергію. Крім того, магній бере участь у синтезі АТФ і стабілізує мембранні структури тилакоїдів, що забезпечує ефективний транспорт електронів. За достатнього забезпечення магнієм зменшується утворення реактивних форм кисню, знижується рівень оксидативного стресу і підтримується висока інтенсивність фотосинтезу навіть за несприятливих умов. У практичних умовах ефективним заходом є внесення сульфату магнію в нормі 5–10 кг/га, що дозволяє швидко компенсувати дефіцит цього елемента і стабілізувати роботу фотосинтетичного апарату.
Не менш важливим елементом є залізо, яке входить до складу ключових компонентів електрон-транспортного ланцюга хлоропластів, зокрема ферредоксину і цитохромів. Воно забезпечує перенесення електронів у процесі фотосинтезу і синтез відновлювальних еквівалентів, необхідних для фіксації вуглецю. За дефіциту заліза ефективність електронного транспорту різко знижується, що призводить до розвитку фотостресу, пошкодження фотосистем і зменшення продуктивності рослин. Забезпечення рослин доступним залізом дозволяє підтримувати стабільну роботу фотосинтетичного апарату і підвищує їх стійкість до стресових факторів.
Особливе значення також має марганець, який є незамінним компонентом фотосистеми II і бере безпосередню участь у процесі фотолізу води — початковому етапі фотосинтезу, під час якого утворюються електрони, протони і молекулярний кисень. Саме цей процес забезпечує функціонування всієї фотосинтетичної електрон-транспортної системи. За достатнього забезпечення марганцем підвищується стабільність фотосистем, зменшується ризик фотопошкодження і підтримується висока ефективність фотосинтезу навіть за умов стресу.
Таким чином, магній, залізо і марганець відіграють фундаментальну роль у підтриманні структурної і функціональної цілісності хлоропластів і забезпеченні стабільного фотосинтезу. Їх своєчасне і збалансоване застосування дозволяє зменшити негативний вплив стресових факторів, знизити рівень оксидативного пошкодження, зберегти енергетичний потенціал рослин і створити фізіологічні передумови для формування високого врожаю навіть за несприятливих умов вирощування.
Мал. 2. Регулятори технології вирощування рослин.
Таблиця 1
Підтримка фотосинтезу як ключового джерела енергії рослини в умовах стресу
| Елемент | Основна фізіологічна роль | Механізм антистресової дії | Норма внесення | Оптимальна фаза застосування | Очікуваний результат |
| Магній (Mg) | Центральний атом молекули хлорофілу; активатор ферментів фотосинтезу | • Знижує утворення ROS • Стабілізує тилакоїдні мембрани • Підтримує активність Rubisco |
5–10 кг/га MgSO₄ (листково або ґрунтово) | Початок активної вегетації Кущення – прапорцевий лист (пшениця) 4–8 листків (соняшник) |
Збереження інтенсивності фотосинтезу Зменшення фотодеструкції хлорофілу |
| Залізо (Fe) | Компонент цитохромів і ферредоксину; транспорт електронів | • Підтримує роботу PSI і PSII • Зменшує фотоінгібування • Підтримує синтез хлорофілу |
0,5–1,5 кг/га (хелати Fe, листково) | Ранній вегетативний ріст Періоди холодового або світлового стресу |
Підтримка електронного транспорту Зменшення хлорозу |
| Марганець (Mn) | Ключовий елемент водорозщеплюючого комплексу PSII | • Забезпечує фотоліз води • Підтримує утворення ATP і NADPH • Знижує оксидативний стрес |
1–3 кг/га MnSO₄ або хелати Mn | Фаза інтенсивного росту листкової поверхні До або під час стресу |
Стабільна робота фотосистеми II Підвищення енергетичного статусу клітини |
Загалом, ключовим фізіологічним принципом виживання і продуктивності рослини є збереження функціональної цілісності хлоропластів як головного джерела енергії клітини. Саме фотосинтетичний апарат забезпечує утворення АТФ, відновлювальних еквівалентів і вуглецевих скелетів, необхідних як для підтримання базового метаболізму, так і для реалізації ростових програм. За дії будь-якого стресового чинника фотосинтез є однією з перших і найбільш чутливих систем, що зазнає пошкодження. Порушення електронного транспорту, накопичення активних форм кисню та деградація хлорофілу швидко призводять до енергетичного дефіциту. У цьому контексті саме ступінь збереження функції хлоропластів визначає, чи зможе рослина вижити, наскільки швидко вона відновиться після стресу і який рівень врожайності буде реалізований у підсумку.
Сучасні фізіолого-біохімічні дослідження переконливо показують, що магній, залізо і марганець формують фундамент стабільності фотосинтетичного апарату в умовах стресу. Магній, як центральний атом молекули хлорофілу, безпосередньо визначає ефективність поглинання світла; залізо забезпечує функціонування компонентів електронтранспортного ланцюга; марганець є критично необхідним для роботи фотосистеми II та фотолізу води. Сукупна дія цих елементів підтримує безперервність електронного транспорту, обмежує утворення активних форм кисню, запобігає деградації пігментів і дозволяє клітині зберігати енергетичний баланс навіть за несприятливих умов.
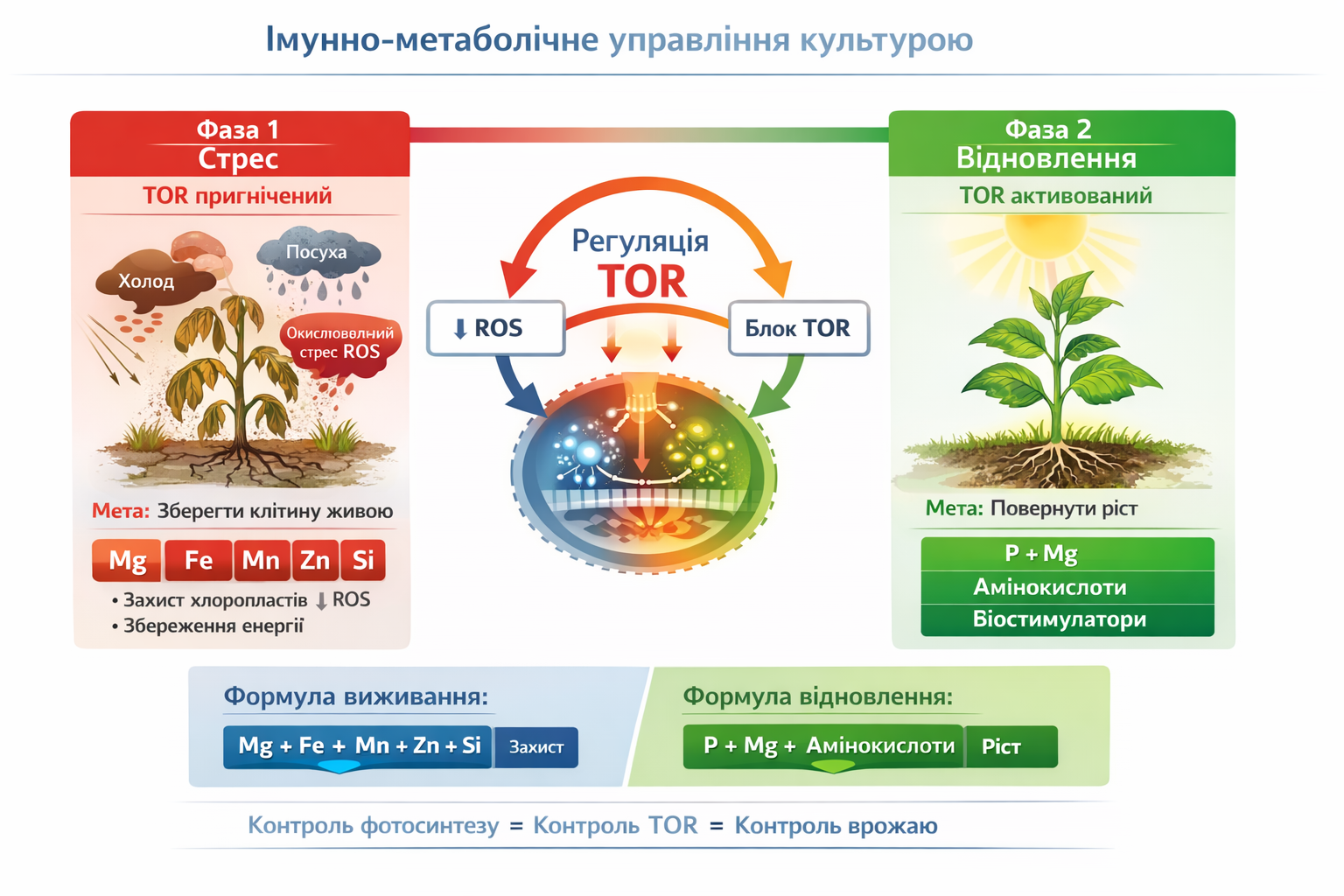
У межах інтегративної моделі імунно-метаболічної регуляції ця закономірність відображається у вигляді двох фундаментальних фізіологічних фаз. Перша фаза — фаза стресу, що характеризується пригніченням TOR-сигналізації та переходом клітини у режим виживання. У цей період головним завданням є не ріст, а збереження життєздатності клітинних структур. Критичну роль відіграють магній, залізо, марганець, цинк і кремній, які стабілізують мембранні системи, знижують рівень оксидативного пошкодження, запобігають руйнуванню хлоропластів і мінімізують енергетичні втрати. Така стабілізація дозволяє клітині зберегти базову метаболічну активність і підготуватися до наступного етапу.
Друга фаза — фаза відновлення, яка настає після послаблення або усунення стресового чинника і пов’язана з реактивацією TOR-сигналізації. У цей період клітина переходить від режиму збереження до режиму продуктивності. Ключового значення набувають фосфор, магній, амінокислоти та регуляторні сполуки, які забезпечують відновлення синтетичних процесів, реактивацію фотосинтезу і запуск програм клітинного росту та диференціації. Саме в цій фазі відбувається компенсація попередніх пошкоджень і реалізація потенціалу врожайності.
Принципово важливим фундаментальним положенням є те, що активація TOR-залежних ростових процесів можлива лише за умови функціонально збережених або відновлених хлоропластів. Якщо ріст активується в умовах пошкодженого фотосинтетичного апарату, клітина опиняється в стані критичного енергетичного дефіциту, що призводить до виснаження ресурсів, втрати продуктивності або навіть загибелі. Таким чином, енергетична готовність клітини є обов’язковою передумовою переходу від виживання до росту.
З практичної точки зору ця закономірність формує чітку фізіологічну логіку управління рослиною. На етапі стресу ключового значення набуває так звана формула виживання, що включає магній, залізо, марганець, цинк і кремній, які забезпечують стабілізацію фотосинтетичного апарату і збереження енергетичного потенціалу. Після стабілізації клітини настає фаза відновлення, де провідну роль відіграє поєднання фосфору, магнію та амінокислот, що забезпечує ефективну реактивацію TOR-сигналізації і повернення до продуктивного росту.
Реалізація такої послідовної стратегії безпосередньо відображається на рівні агроценозу, оскільки забезпечує значно швидше відновлення рослин після стресових подій, підвищує інтенсивність фотосинтезу і сприяє зростанню врожайності. У своїй основі цей підхід відображає універсальний біологічний принцип: контроль фотосинтетичного апарату означає контроль енергетичного стану клітини; контроль енергії визначає активність TOR-сигналізації; а контроль TOR, у свою чергу, визначає реалізацію росту і формування врожаю.
Як керувати ROS — головним фактором пошкодження?
Регуляція рівня реактивних форм кисню (ROS) є одним із ключових аспектів забезпечення стійкості рослин до стресових факторів і збереження їх продуктивності. У нормальних фізіологічних умовах ROS утворюються в клітинах як побічні продукти фотосинтезу і дихання та виконують сигнальні функції, беручи участь у регуляції росту, розвитку і захисних реакцій. Однак за дії стресів, таких як посуха, температурні екстремуми, гербіцидне навантаження або дефіцит елементів живлення, їх утворення різко зростає, що призводить до розвитку оксидативного стресу. Надлишкове накопичення ROS викликає пошкодження клітинних мембран, білків, ліпідів і нуклеїнових кислот, порушує роботу хлоропластів і мітохондрій, пригнічує фотосинтез і синтез білка, що в кінцевому результаті проявляється у вигляді пригнічення росту, зниження продуктивності і навіть загибелі клітин.
Ефективним шляхом мінімізації негативного впливу ROS є застосування сполук, які виконують функції антиоксидантів або стабілізаторів клітинного метаболізму. Однією з найбільш ефективних речовин цього типу є гліцин-бетаїн — низькомолекулярна органічна сполука, яка виконує роль осмопротектора і стабілізатора білків та мембран. Гліцин-бетаїн забезпечує захист фотосинтетичного апарату, зокрема хлоропластів, підтримує стабільність фотосистем і ферментних комплексів та значно зменшує рівень утворення реактивних форм кисню. Завдяки цьому зберігається ефективність фотосинтезу і підтримується енергетичний баланс клітини навіть в умовах стресу.
Важливу роль у захисті клітин також відіграє амінокислота пролін, яка накопичується в рослинах у відповідь на дію несприятливих факторів. Пролін виконує кілька функцій: він стабілізує структуру білків і мембран, підтримує осмотичний баланс і одночасно виступає резервним джерелом енергії та відновлювальних еквівалентів. Після припинення дії стресу пролін може швидко включатися в метаболізм, забезпечуючи енергетичну підтримку процесів відновлення і росту.
Особливе місце серед антиоксидантних сполук займає мелатонін, який вважається одним із найпотужніших природних антиоксидантів у рослинних клітинах. Він безпосередньо нейтралізує реактивні форми кисню, активує антиоксидантні ферментні системи, включаючи супероксиддисмутазу, каталазу і пероксидази, та стабілізує роботу хлоропластів і мітохондрій. Завдяки цьому мелатонін значно підвищує стійкість рослин до широкого спектра стресових факторів, зокрема посухи, низьких температур і гербіцидного навантаження, зменшуючи рівень клітинних пошкоджень і сприяючи збереженню фізіологічної активності.
Таким чином, управління рівнем реактивних форм кисню є критично важливим елементом сучасних технологій вирощування рослин. Застосування таких сполук, як гліцин-бетаїн, пролін і мелатонін, дозволяє ефективно контролювати оксидативний стрес, захищати фотосинтетичний апарат і клітинні структури, підтримувати енергетичний баланс і забезпечувати стабільний ріст і розвиток рослин навіть за умов дії несприятливих факторів. Це створює фізіологічну основу для збереження високої продуктивності і реалізації генетичного потенціалу врожайності.
Таблиця 2
Регуляція активних форм кисню (ROS) як ключовий механізм
збереження продуктивності рослин
| Антистресова речовина | Біохімічний механізм дії | Вплив на ROS-гомеостаз | Клітинний і фізіологічний ефект | Практичне значення у полі | Очікуваний вплив на врожайність, % |
| Гліцин-бетаїн | Осмопротектор; стабілізує білки та ліпідні мембрани хлоропластів; підтримує PSII | Значно знижує утворення ROS у хлоропластах; захищає фотосинтетичний електронний транспорт | Захист фотосистем; збереження структури хлоропластів; підтримка фотосинтезу | Особливо ефективний при посусі, високих температурах, гербіцидному стресі | +10–25 % |
| Пролін | Осморегулятор; виконує роль резервного джерела енергії і відновлювальних еквівалентів | Буферизація ROS; стабілізація редокс-балансу клітини | Захист мембран; підтримка метаболізму; прискорення відновлення після стресу | Важливий при водному дефіциті, холодовому та сольовому стресі | +8–20 % |
| Мелатонін | Потужний універсальний антиоксидант; регулятор експресії антиоксидантних ферментів (SOD, CAT, APX) | Безпосередньо нейтралізує ROS і активує антиоксидантну систему | Захист мітохондрій і хлоропластів; збереження клітинної життєздатності | Найефективніший при комплексних стресах: посуха, холод, гербіциди, температурні коливання | +15–30 % |
| Аскорбат (вітамін C) | Ключовий компонент аскорбат-глутатіонового циклу | Детоксикація H₂O₂; підтримка редокс-гомеостазу | Захист фотосинтетичного апарату; стабілізація метаболізму | Ефективний у період інтенсивного світлового і температурного стресу | +8–18 % |
| Глутатіон | Центральний редокс-буфер клітини; регуляція сигнальних шляхів стресу | Нейтралізація перекисів; контроль окисно-відновного стану | Підвищення стресостійкості; збереження функції ферментів | Важливий при гербіцидному та оксидативному стресі | +10–20 % |
У цілому, ключовим науково-практичним висновком сучасної фізіології рослин є розуміння того, що активні форми кисню (ROS) виступають центральним регулятором, який визначає межу між оборотним стресом і незворотним пошкодженням клітини. У помірних концентраціях ROS виконують сигнальну функцію, активуючи захисні механізми, однак їх надмірне накопичення призводить до окисного руйнування ліпідів мембран, денатурації білків і деградації хлоропластів. Саме пошкодження фотосинтетичного апарату є критичним моментом, оскільки фотосинтез є основним джерелом енергії для підтримання життєздатності клітини та формування врожаю. Пригнічення фотосинтезу запускає каскад енергетичного дефіциту, який обмежує процеси відновлення, росту та продуктивності, що в кінцевому результаті проявляється у прямих втратах урожайності.
Найефективніша стратегія захисту рослин у цих умовах базується на використанні сполук із подвійним механізмом дії, які одночасно забезпечують безпосередню нейтралізацію надлишкових активних форм кисню та стабілізують структурну і функціональну цілісність енергетичного і фотосинтетичного апарату клітини. Такий підхід дозволяє не лише зменшити рівень окисного пошкодження, але й зберегти здатність рослини до швидкого відновлення після припинення дії стресового фактора.
У практичному вимірі сучасних агротехнологій сформувалася базова формула ефективного контролю ROS, яка включає комплекс ключових антистресових метаболітів — гліцин-бетаїн, пролін і мелатонін. Гліцин-бетаїн виконує функцію потужного осмопротектора, стабілізуючи мембрани та білкові комплекси хлоропластів і безпосередньо захищаючи фотосистеми від окисного руйнування. Пролін, у свою чергу, виступає універсальним клітинним протектором і резервним джерелом енергії, підтримуючи редокс-баланс і сприяючи відновленню метаболічних процесів. Особливе місце займає мелатонін, який є одним із найпотужніших природних антиоксидантів, здатних як безпосередньо нейтралізувати ROS, так і активувати внутрішні антиоксидантні системи клітини.
Фізіологічним результатом застосування такого підходу є суттєве зниження рівня оксидативного пошкодження клітинних структур, яке може зменшуватися на 30–60 %, збереження активності фотосинтетичного апарату на рівні, що перевищує контрольні рослини на 15–35 %, а також значне прискорення процесів відновлення після дії стресових факторів — на 20–40 %. У виробничих умовах це безпосередньо трансформується у збереження та додатковий приріст врожайності, який зазвичай становить 10–30 % залежно від культури та інтенсивності стресу.
Таким чином, фундаментальний стратегічний принцип сучасного управління продуктивністю рослин полягає у тому, що контроль рівня активних форм кисню є основою збереження функціональної цілісності хлоропластів, а отже — збереження енергетичного потенціалу клітини. Саме підтримання енергетичного балансу визначає здатність рослини не лише вижити у стресових умовах, але й повністю реалізувати свій генетично закладений потенціал продуктивності. У цьому контексті контроль ROS фактично означає контроль енергії, а контроль енергії — це контроль кінцевого врожаю.
Як керувати амінокислотами?
Регуляція амінокислотного живлення є одним із найбільш ефективних інструментів управління стійкістю рослин до стресу і їх здатністю до швидкого відновлення та подальшого росту. Амінокислоти виконують не лише функцію структурних компонентів білків, але й відіграють ключову роль як сигнальні молекули, регулятори метаболізму і джерела енергії. В умовах стресу їх значення особливо зростає, оскільки синтез власних амінокислот у рослині потребує значних витрат енергії, яка в цей період є обмеженим ресурсом. Саме тому екзогенне внесення амінокислот дозволяє зменшити енергетичні витрати клітини, підтримати метаболічну стабільність і забезпечити більш ефективну адаптацію до несприятливих умов.
Ключовим практичним питанням є визначення оптимального часу застосування амінокислот. Найбільш ефективною є стратегія їх використання як до настання стресу, так і після його дії. Внесення амінокислот до дії стресового фактора сприяє активації захисних механізмів, стабілізації клітинних структур і підвищенню загальної фізіологічної готовності рослини до несприятливих умов. У цей період амінокислоти виконують функцію метаболічних сигналів, які стимулюють синтез антиоксидантних ферментів, осмопротекторів і інших захисних сполук. Після припинення дії стресу їх роль змінюється — вони стають важливим фактором відновлення ростових процесів, забезпечуючи клітини готовими субстратами для синтезу білків, ферментів і структурних компонентів, а також сприяючи активації енергетичного метаболізму.
Особливе значення мають окремі амінокислоти, які виконують специфічні функції у формуванні стресостійкості і підтриманні фотосинтезу. Зокрема, пролін є однією з основних антистресових амінокислот, яка накопичується у відповідь на дію несприятливих факторів і забезпечує стабілізацію білків і мембран, а також виконує функцію резервного джерела енергії. Глутамат займає центральне місце у метаболізмі азоту і вуглецю, забезпечуючи клітину енергією і виступаючи попередником для синтезу інших амінокислот і важливих метаболітів. Аргінін є ключовим попередником поліамінів — сполук, які беруть участь у регуляції клітинного поділу, стабілізації мембран і захисті від оксидативного стресу. Гліцин відіграє важливу роль у функціонуванні фотосинтетичного апарату, оскільки входить до складу ферментів фотодихання і бере участь у синтезі хлорофілу, що безпосередньо впливає на ефективність фотосинтезу.
Таким чином, цілеспрямоване застосування амінокислот дозволяє ефективно керувати фізіологічним станом рослин у різні періоди їх розвитку і адаптації до стресу. Використання амінокислот до дії несприятливих факторів підвищує рівень захисту і стійкості, тоді як їх застосування після стресу прискорює відновлення метаболізму, стимулює синтез білка і активує ростові процеси. Такий підхід забезпечує збереження фотосинтетичної активності, оптимізацію енергетичного балансу і створює фізіологічні передумови для формування високого рівня продуктивності навіть в умовах стресового навантаження.
Таблиця 3
Функціональна роль амінокислот у регуляції стресостійкості та росту рослин
| Амінокислота | Основна фізіологічна функція | Молекулярний механізм дії | Вплив на SnRK1–TOR баланс | Оптимальна фаза застосування | Вплив на фотосинтез | Вплив на стресостійкість | Очікуваний вплив на врожайність (%) |
| Пролін | Антистресовий осмопротектор | Стабілізує білки, мембрани та ферменти, нейтралізує ROS | Активує SnRK1-залежний захист | До і під час стресу | Захищає фотосистеми | Значно підвищує | +10–25 % |
| Глутамат | Центральний енергетичний метаболіт | Донор азоту і вуглецю для циклу Кребса та синтезу амінокислот | Сприяє реактивації TOR | Після стресу | Прискорює відновлення | Помірно підвищує | +10–20 % |
| Аргінін | Попередник поліамінів | Субстрат синтезу поліамінів (путресцин, спермідин), регулює поділ клітин | Сприяє активації TOR | До і після стресу | Покращує стабільність хлоропластів | Підвищує | +10–20 % |
| Гліцин | Компонент фотосинтетичного метаболізму | Учасник фотодихання і синтезу хлорофілу | Сприяє переходу SnRK1 → TOR | Після стресу | Значно покращує | Помірно підвищує | +10–25 % |
Таблиця 4
Підхід щодо застосування амінокислот
| Фаза | Мета | Роль амінокислот | Основний ефект |
| До стресу | Підготовка клітини | Активація захисних механізмів | Підвищення стійкості |
| Під час стресу | Захист клітин | Стабілізація білків і мембран | Збереження життєздатності |
| Після стресу | Відновлення росту | Активація TOR і синтезу білка | Відновлення врожайності |
Загалом, ключовий фізіологічний принцип застосування амінокислот полягає в тому, що вони виконують подвійну, стратегічно важливу функцію у регуляції стану рослини залежно від її метаболічного режиму. У фазі стресу, коли домінує енергозберігаюча SnRK1-регуляція, амінокислоти виступають як фактори виживання: вони стабілізують білкові структури, підтримують осмотичний баланс, зменшують оксидативне пошкодження та забезпечують клітину доступними метаболічними субстратами, необхідними для підтримання базової життєздатності. У цей період їхня роль полягає насамперед у збереженні функціональної цілісності клітини та запобіганні незворотним пошкодженням. Натомість у фазі відновлення, коли стресовий вплив зменшується і реактивується TOR-залежний режим, амінокислоти виконують іншу, не менш важливу функцію — вони стають джерелом швидкодоступного азоту, енергії та сигнальних молекул, необхідних для реактивації фотосинтезу, синтезу білків, поділу клітин і формування біомаси. Саме в цій фазі вони безпосередньо впливають на реалізацію врожайного потенціалу.
Найважливіший практичний висновок полягає в тому, що максимальна ефективність амінокислот визначається не частотою або регулярністю їх внесення, а точністю вибору фізіологічного моменту застосування. Їх використання має бути синхронізоване з критичними фазами стану рослини: до настання або на початку стресу амінокислоти доцільно застосовувати для активації захисних механізмів, стабілізації клітинних структур і підвищення стійкості, тоді як після послаблення або припинення стресового впливу їх внесення забезпечує ефективне переключення метаболізму у TOR-залежний режим росту, прискорює відновлення фотосинтетичної активності та сприяє формуванню врожаю. Таким чином, амінокислоти є не просто джерелом живлення, а інструментом регуляції переходу між режимами виживання і продуктивності, і саме правильний момент їх застосування визначає кінцевий агрономічний результат.

Мал. 3. Гербіцидний стрес.
Як керувати GABA?
Гамма-аміномасляна кислота (GABA) є одним із ключових антистресових метаболітів у рослин, який виконує важливу роль у регуляції їх адаптації до несприятливих умов середовища (посухи, засолення, гербіцидів). Препарати з амінокислотами стимулюють синтез GABA. Вона швидко накопичується у клітинах у відповідь на дію різних стресових факторів і виступає не лише як проміжний продукт метаболізму, але і як сигнальна молекула, що координує фізіологічні та біохімічні захисні реакції. Основною функцією GABA є стабілізація клітинного метаболізму в умовах енергетичного дефіциту і оксидативного стресу. Вона сприяє підтриманню роботи мітохондрій, забезпечує альтернативний шлях утворення енергії через так званий GABA-шунт і одночасно зменшує накопичення реактивних форм кисню, які є головною причиною пошкодження клітин при стресі.
Завдяки цим властивостям GABA значно підвищує стійкість рослин до широкого спектра абіотичних факторів, зокрема посухи, засолення ґрунту і гербіцидного навантаження. В умовах посухи вона допомагає підтримувати водний баланс клітини і зберігати функціонування фотосинтетичного апарату. При сольовому стресі GABA сприяє регуляції іонного гомеостазу і зменшує токсичний вплив надлишку натрію. У випадку гербіцидного стресу вона підтримує енергетичний метаболізм і прискорює процеси відновлення, що дозволяє рослині швидше повернутися до нормального росту.
З практичної точки зору, важливою особливістю є те, що синтез GABA тісно пов’язаний із метаболізмом амінокислот, насамперед глутамату, який є її безпосереднім попередником. Саме тому застосування препаратів, що містять амінокислоти, є ефективним способом стимуляції її утворення в рослинах. Такі препарати забезпечують клітини готовими субстратами для синтезу GABA, що дозволяє швидко активувати захисні механізми і підвищити стійкість рослин до стресу. Крім того, це сприяє стабілізації енергетичного балансу, збереженню функціональної активності хлоропластів і мітохондрій та прискорює відновлення фізіологічних процесів після припинення дії несприятливих факторів.
Таким чином, управління рівнем GABA через систему живлення, зокрема шляхом застосування амінокислот, є ефективним інструментом підвищення стресостійкості рослин. Це дозволяє зменшити ступінь пошкодження клітин, підтримати енергетичний метаболізм і забезпечити більш стабільний ріст і розвиток навіть у складних умовах вирощування, що в кінцевому результаті сприяє збереженню і підвищенню врожайності.
Таблиця 5
Регуляція стресостійкості рослин через стимуляцію синтезу GABA
| Параметр | Опис / Дія | Практичне застосування |
| Метаболіт | GABA (γ-аміномасляна кислота) | Один із ключових антистресових метаболітів рослини |
| Фізіологічний ефект | Підвищує загальну стійкість до абіотичних і хімічних стресів | • Посуха • Засолення • Гербіцидне навантаження |
| Регуляторний механізм | Знижує рівень оксидативного стресу, стабілізує клітинний редокс-стан, підтримує енергетичний баланс через GABA-шунт | Синтез активується накопиченням глутамату та екзогенними амінокислотами |
| Практичне втручання | Застосування амінокислотних препаратів (позакоренево або через ґрунт) | Стимулює внутрішній синтез GABA, підвищує адаптивну реакцію та стабільність метаболізму |
Як керувати енергією рослини?
Енергетичний баланс є фундаментальною основою всіх фізіологічних процесів у рослині, оскільки саме енергія визначає здатність клітин підтримувати ріст, фотосинтез, транспорт речовин і формування врожаю. В умовах стресу, таких як посуха, температурні коливання, гербіцидне навантаження або дефіцит елементів живлення, рослина часто стикається з явищем енергетичного голодування, коли утворення енергії зменшується, а її витрати на підтримання життєво важливих процесів і захисних реакцій, навпаки, різко зростають. У таких умовах порушується синтез АТФ — універсального енергетичного носія клітини, що призводить до пригнічення росту, уповільнення метаболізму і зниження здатності рослини відновлюватися після стресу. Тому одним із ключових завдань сучасної системи живлення є забезпечення рослин елементами, які безпосередньо беруть участь у процесах утворення, накопичення і використання енергії.
Особливу роль у цьому процесі відіграє фосфор, який є структурним компонентом молекули АТФ і бере участь у всіх енергетичних реакціях клітини. Він забезпечує накопичення і перенесення енергії, необхідної для синтезу органічних речовин, функціонування мембранних транспортних систем і роботи ферментів. За достатнього забезпечення фосфором підтримується висока інтенсивність енергетичного обміну, активний синтез білка і стабільна робота фотосинтетичного апарату. Особливо важливим є його доступність у періоди активного росту і після дії стресу, коли потреба в енергії різко зростає.
Не менш важливим елементом енергетичного метаболізму є магній, який бере безпосередню участь у синтезі АТФ і функціонуванні ферментів, що забезпечують енергетичні перетворення. Магній стабілізує структуру молекули АТФ, забезпечуючи її біологічну активність, а також відіграє ключову роль у роботі хлоропластів, де відбувається утворення енергії під час фотосинтезу. Крім того, він сприяє підтриманню структурної цілісності мембран і оптимальному функціонуванню ферментних систем, що дозволяє рослині ефективніше використовувати наявні енергетичні ресурси.
Таким чином, забезпечення рослин фосфором і магнієм є одним із найважливіших практичних заходів для запобігання енергетичному голодуванню і підтримання високої метаболічної активності. Їх своєчасне і збалансоване застосування дозволяє зберегти ефективність фотосинтезу, забезпечити стабільний синтез АТФ, підтримати процеси росту і відновлення після стресу та створити фізіологічні передумови для формування високого рівня продуктивності рослин.
Як правильно використовувати антистресанти?
Правильне використання антистресантів є одним із ключових інструментів управління фізіологічною стійкістю рослин і збереження їх продуктивності в умовах дії несприятливих факторів. Основним практичним питанням при цьому є визначення оптимального часу їх застосування, оскільки саме від цього залежить ефективність їх дії і кінцевий результат. Антистресові препарати не є лише засобами відновлення — їх головна цінність полягає у здатності підготувати рослину до стресу, активувати її власні захисні механізми і мінімізувати ступінь пошкодження клітинних структур.
Найвища ефективність антистресантів досягається у випадку їх застосування до настання стресового фактору. У цей період вони виконують функцію метаболічних активаторів, які стимулюють синтез захисних сполук, таких як антиоксиданти, осмопротектори, поліаміни та інші компоненти системи стресостійкості. Це дозволяє стабілізувати клітинні мембрани, підвищити ефективність роботи фотосинтетичного апарату, зменшити накопичення реактивних форм кисню і забезпечити достатній рівень енергетичного забезпечення клітини. У результаті рослина входить у стресовий період у фізіологічно підготовленому стані, що дозволяє значно зменшити негативні наслідки. За такого підходу ефективність антистресантів може становити 70–90 %, оскільки вони працюють як інструмент профілактики, а не лише як засіб лікування.
У випадку застосування антистресантів після дії стресу їх функція змінюється — вони вже не можуть повністю запобігти пошкодженням, а лише сприяють відновленню метаболічних процесів і регенерації клітин. У цей період антистресові речовини допомагають стабілізувати роботу хлоропластів і мітохондрій, активувати синтез білка, зменшити рівень оксидативного стресу і прискорити відновлення росту. Однак частина клітинних структур до цього моменту вже може бути пошкоджена, а частина енергетичних ресурсів втрачена, тому загальна ефективність такого застосування значно нижча і зазвичай становить лише 30–50 %. Це пояснюється тим, що відновлення завжди потребує більше часу і ресурсів, ніж попередження пошкодження.
Таким чином, найбільш ефективною стратегією використання антистресантів є їх профілактичне застосування до настання стресових умов. Це дозволяє активувати природні механізми стійкості рослин, зберегти функціонування фотосинтетичного апарату, запобігти енергетичному дефіциту і мінімізувати пошкодження клітин. Застосування антистресантів після стресу також має важливе значення, однак його слід розглядати насамперед як інструмент відновлення, а не повноцінної профілактики. Раціональне поєднання профілактичного і відновлювального застосування антистресових препаратів дозволяє забезпечити стабільний фізіологічний стан рослин і створює оптимальні умови для реалізації їх продуктивного потенціалу.
Таблиця 6
Правила використання антистресантів
| Параметр | Рекомендація | Практичний ефект / Ефективність |
| Ключове питання | Коли застосовувати антистресант? | Максимальна користь досягається при правильному фазуванні внесення |
| Правило №1 | До стресу | 70–90 % ефективності, активує захисні механізми рослини (SnRK1-фаза) |
| Правило №2 | Після стресу | 30–50 % ефективності, підтримка відновлення росту та TOR-фази |
Чим обробити рослину ефективно при абіотичних стресах ?
Одним із найважливіших практичних питань сучасної технології вирощування сільськогосподарських культур є правильний вибір фізіологічно обґрунтованих заходів для захисту рослин у періоди дії основних абіотичних і хімічних стресових факторів, зокрема високих температур, посухи, засоленості та кислотності ґрунту, низьких температур і заморозків, а також гербіцидного навантаження. Усі ці фактори мають спільну фізіологічну основу впливу — вони викликають порушення енергетичного метаболізму, пригнічення фотосинтезу, накопичення реактивних форм кисню і дестабілізацію клітинних структур. Тому головним завданням системи антистресового захисту є стабілізація водного балансу, підтримання енергетичного забезпечення клітини, захист фотосинтетичного апарату і активація власних механізмів адаптації рослини.
В умовах високих температур, посухи, засоленості і підвищеної кислотності ґрунту найбільш ефективними заходами є застосування калію, амінокислот і гліцин-бетаїну. Калій відіграє ключову роль у регуляції осмотичного тиску і роботи продихів, що дозволяє рослині ефективніше утримувати воду, зменшувати її втрати через транспірацію і підтримувати нормальне функціонування клітин навіть за умов водного дефіциту. Амінокислоти виконують функцію метаболічної підтримки, забезпечуючи клітини готовими субстратами для синтезу білків, ферментів і захисних сполук, а також зменшують енергетичні витрати на відновлення. Особливу ефективність у цих умовах проявляє гліцин-бетаїн, який стабілізує мембрани, захищає фотосинтетичний апарат, знижує рівень оксидативного стресу і забезпечує збереження функціональної активності хлоропластів.
За дії низьких температур і заморозків ключове значення має забезпечення клітини енергією і стабілізація метаболічних процесів. Найбільш ефективними в цьому випадку є фосфор, амінокислоти і мелатонін. Фосфор забезпечує синтез і накопичення АТФ, що дозволяє підтримувати життєво важливі процеси навіть за умов пригнічення фотосинтезу. Амінокислоти стабілізують білкові структури і мембрани, а також виконують роль резервного джерела енергії. Мелатонін, як потужний природний антиоксидант, ефективно нейтралізує реактивні форми кисню, які інтенсивно утворюються при температурному стресі, і захищає клітинні органели від пошкодження, підвищуючи загальну стійкість рослин до холоду.
Особливу категорію становить гербіцидний стрес, який безпосередньо пов’язаний із порушенням метаболізму, пригніченням фотосинтезу і накопиченням токсичних продуктів у клітині. У таких умовах найбільш ефективним є застосування амінокислот, магнію і антиоксидантів. Амінокислоти прискорюють відновлення синтезу білка і ферментних систем, магній стабілізує структуру хлорофілу і забезпечує відновлення фотосинтетичної активності, а антиоксиданти знижують рівень оксидативного пошкодження клітинних структур.
Таким чином, правильний вибір антистресових компонентів залежно від типу стресового фактору дозволяє ефективно управляти фізіологічним станом рослин. Використання калію, амінокислот і гліцин-бетаїну при посусі і високих температурах, фосфору, амінокислот і мелатоніну при холодовому стресі, а також амінокислот, магнію і антиоксидантів після гербіцидного впливу забезпечує стабілізацію метаболізму, збереження фотосинтетичної активності і створює оптимальні умови для відновлення росту і формування високого врожаю навіть у складних умовах вирощування.
Таблиця 7
Вибір антистресових компонентів залежно від типу стресового фактору
| Практичне питання | Рекомендації / Засоби | Примітки / Ефект |
| Чим обробити рослину при посусі | Калій, амінокислоти, гліцин-бетаїн | Підвищує стійкість до водного дефіциту |
| Чим обробити рослину при холоді | Фосфор, амінокислоти, мелатонін | Підтримка енергетичного та гормонального балансу |
| Чим обробити рослину при гербіцидному стресі | Амінокислоти, магній, антиоксиданти | Захист клітин та фотосинтезу, зменшення ROS |
| Як швидко відновити рослину після стресу | Амінокислоти, магній, азот | Активує TOR-фазу росту та фотосинтез |
| Як підвищити врожайність при стресі | Кремній, калій, амінокислоти | Оптимізація росту, стабілізація клітинних структур |
| Найцінніша практична відповідь | Стрес = енергетичний дефіцит; Зменшити втрати енергії: антиоксиданти; Збільшити виробництво енергії: Mg, Fe, P | Базова логіка управління енергією в рослині |
| Найважливіше практичне правило | До стресу: антистресанти; Після стресу: добрива для росту | Фазове управління SnRK1 → TOR |
Як швидко відновити рослину після стресу найефективніше?
Одним із найважливіших прикладних завдань сучасної технології вирощування є максимально швидке і повноцінне відновлення рослини після перенесеного стресу, оскільки саме період після стресу визначає майбутню продуктивність, інтенсивність росту та рівень врожайності. У цей час рослина перебуває у стані енергетичного дефіциту, пошкодженого метаболізму та пригніченого синтезу структурних і функціональних компонентів клітини. Найефективнішим інструментом відновлення є застосування амінокислот, які виконують роль готових структурних блоків для синтезу білків, ферментів і регуляторних молекул, що дозволяє рослині економити енергію і значно швидше відновлювати фізіологічні процеси.
Не менш важливу роль відіграє магній, який є центральним елементом молекули хлорофілу та критично необхідний для відновлення фотосинтезу і синтезу АТФ, тобто енергетичної валюти клітини. Забезпечення рослини доступним азотом також має ключове значення, оскільки він необхідний для синтезу нових білків, ферментів і нуклеїнових кислот, що забезпечують відновлення росту і формування нових тканин. Комплексне застосування амінокислот, магнію та азоту дозволяє максимально швидко відновити фотосинтетичну активність, енергетичний баланс і метаболічну активність рослини, що є основою повернення до інтенсивного росту і формування високого рівня продуктивності навіть після сильних стресових впливів.
Як підвищити врожайність при стресі найефективніше?
Одним із найважливіших прикладних завдань технології вирощування є підвищення врожайності рослин в умовах стресу, оскільки саме стресові фактори — посуха, високі температури, засолення, дія гербіцидів чи низькі температури — є головною причиною втрати продуктивності. Найефективнішою стратегією у таких умовах є застосування речовин, які одночасно підвищують фізіологічну стійкість рослини та підтримують її метаболічну активність. Особливу роль відіграє кремній, який зміцнює клітинні стінки, зменшує втрати води, підвищує механічну міцність тканин і знижує проникнення токсичних сполук, що в результаті значно підвищує загальну стійкість рослини до несприятливих факторів.
Не менш важливим є калій, який регулює водний баланс, контролює роботу продихів, покращує транспорт асимілянтів і підвищує ефективність фотосинтезу, що особливо критично в умовах посухи та високих температур. Водночас застосування амінокислот забезпечує рослину готовими метаболічними компонентами, які прискорюють синтез білків, ферментів і захисних молекул, знижують енергетичні витрати на відновлення і дозволяють підтримувати ріст навіть у несприятливих умовах. Комплексне використання кремнію, калію та амінокислот дозволяє зберегти активність фотосинтезу, стабілізувати водний режим, захистити клітинні структури і забезпечити ефективне формування врожаю навіть за наявності сильного стресового навантаження.
Стрес = енергетичний дефіцит.
Найцінніша практична відповідь і ключовий принцип сучасного управління рослиною у стресових умовах базується на розумінні того, що стрес — це насамперед стан енергетичного дефіциту. Всі основні стресові фактори — посуха, високі чи низькі температури, засолення, гербіциди — призводять до порушення роботи фотосинтезу, дихання та метаболізму, що зменшує енергетичні ресурси клітини і, як наслідок, пригнічує ріст, відновлення та формування врожаю. Тому головне завдання полягає у двох взаємопов’язаних підходах: або зменшити втрати енергії, або максимально підвищити її виробництво.
Зменшення енергетичних витрат досягається через застосування антиоксидантів, які нейтралізують реактивні форми кисню, стабілізують клітинні мембрани та органели, зменшують оксидативне пошкодження і дозволяють клітині ефективніше використовувати наявну енергію. Таким чином, антиоксиданти виступають як щит, що захищає фотосинтетичний апарат і метаболічні системи, запобігаючи зайвим витратам енергії на відновлення.
Паралельно з цим необхідно підвищувати виробництво енергії шляхом забезпечення рослини ключовими елементами метаболізму. Магній, як центральний атом молекули хлорофілу, забезпечує ефективний фотосинтез і синтез АТФ, залізо бере участь у перенесенні електронів у фотосистемах і дихальних ланцюгах, а фосфор є безпосереднім компонентом АТФ і всіх реакцій енергетичного обміну. Комплексне забезпечення рослини цими елементами дозволяє відновити енергетичний баланс, підтримати роботу фотосинтетичного апарату, активувати синтез білків і ферментів та забезпечити стійкість до стресу.
Таким чином, ефективне управління стресом у рослин полягає у збалансованому поєднанні захисту енергії за допомогою антиоксидантів і стимуляції її виробництва через магній, залізо та фосфор. Це створює фізіологічну основу для збереження росту, відновлення після стресу і формування стабільного, високого врожаю навіть за несприятливих умов вирощування.
Найважливіше практичне правило (золотий стандарт) для контролю стресу.
Найважливіше практичне правило, яке можна вважати «золотим стандартом» сучасного управління рослиною в стресових умовах, базується на чіткій стратегії застосування антистресових засобів та добрив у залежності від фаз впливу несприятливих факторів. До настання стресу найефективнішим заходом є використання антистресантів, які активують природні механізми захисту рослини, стабілізують мембрани та органели, зменшують накопичення реактивних форм кисню і забезпечують енергетичну готовність клітини до несприятливих умов. Такий підхід дозволяє максимально знизити пошкодження тканин і зберегти фізіологічну активність рослини під час стресу.
Після завершення дії стресу пріоритетом стає стимуляція ростових процесів і відновлення метаболізму, для чого застосовують добрива, що забезпечують рослину елементами живлення, необхідними для синтезу білків, ферментів, АТФ і фотосинтетичних компонентів. Це дозволяє швидко повернути фотосинтетичну активність, відновити енергетичний баланс і активізувати ріст, формування нових пагонів і генеративних органів. Дотримання цього золотого стандарту — профілактичне застосування антистресантів до стресу і живлення для росту після нього — забезпечує оптимальну фізіологічну готовність рослини, підвищує її стійкість і дозволяє реалізувати максимальний продуктивний потенціал навіть за умов несприятливого середовища.
Таблиця 8
Система антистресового живлення рослин (практичний протокол)
- ДО СТРЕСУ (профілактика)
Мета: активація захисних систем, стабілізація хлоропластів, підготовка до ROS
| Компонент | Норма | Форма | Функція | Практичний ефект |
| Калій (K₂O) | 1–3 кг/га | сульфат, цитрат | осморегуляція | посуха, холод |
| Магній (Mg) | 0,5–1 кг/га | MgSO₄, хелат | стабілізація хлорофілу | збереження фотосинтезу |
| Амінокислоти | 0,5–1,5 л/га | L-форма | синтез антистресових білків | універсальний захист |
| Гліцин-бетаїн | 0,3–0,5 кг/га | позакоренево | захист хлоропластів | посуха, температура |
| Кремній | 0,5–1 л/га | орто-кремнієва кислота | зміцнення клітин | посуха, гербіциди |
| Фосфор | 0,5–1 кг/га | фосфіт, ортофосфат | енергетичний резерв | всі стреси |
- ПІД ЧАС СТРЕСУ (фаза пошкодження)
Мета: зменшення ROS, збереження енергії
| Компонент | Норма | Функція | Ефект |
| Амінокислоти | 1–2 л/га | джерело енергії | швидка стабілізація |
| Магній | 1 кг/га | підтримка фотосинтезу | критично важливо |
| Гліцин-бетаїн | 0,5 кг/га | стабілізація мембран | зменшення пошкоджень |
| Калій | 1–2 кг/га | осмозахист | посуха |
| Антиоксиданти | за регламентом | зниження ROS | захист клітин |
- ПІСЛЯ СТРЕСУ (фаза відновлення)
Мета: активація TOR → стимуляція росту
| Компонент | Норма | Функція | Ефект |
| Азот | 3–10 кг/га | синтез білка | відновлення росту |
| Амінокислоти | 1–2 л/га | відновлення клітин | швидкий ріст |
| Магній | 0,5–1 кг/га | фотосинтез | відновлення |
| Фосфор | 0,5–1 кг/га | ATP | енергія |
| Цитокініни | за регламентом | стимуляція поділу клітин | регенерація |
Спеціалізовані схеми під конкретний стрес.
Спеціалізовані схеми під конкретний стрес дозволяють максимально ефективно підтримати фізіологічний стан рослини та мінімізувати негативні наслідки дії абіотичних і хімічних факторів. У випадку посухи, високих температур, засоленості та підвищеної кислотності ґрунту найефективнішою є схема застосування препаратів за 3–5 днів до настання стресу. Вона включає внесення калію у нормі 2 кг/га для підтримки водного балансу та роботи продихів, амінокислот у кількості 1 л/га для забезпечення клітин готовими субстратами для синтезу білків і ферментів, а також гліцин-бетаїну 0,5 кг/га, який стабілізує мембрани, захищає фотосинтетичний апарат і зменшує оксидативне пошкодження.
При гербіцидному стресі найбільш ефективним є обробіток через 24–48 годин після застосування гербіциду. У цьому випадку рослині необхідна підтримка метаболізму та фотосинтезу, тому рекомендується внесення амінокислот у кількості 1,5 л/га для швидкого відновлення білкового обміну, магнію 1 кг/га для стабілізації хлорофілу і відновлення фотосинтетичної активності, а також фосфору 0,5 кг/га для забезпечення енергетичного балансу та синтезу АТФ.
У разі дії холодового стресу обробку слід проводити за 1–2 дні до настання низьких температур або заморозків. Оптимальна схема включає амінокислоти 1 л/га для підтримки метаболічної активності, фосфор 1 кг/га для забезпечення синтезу енергії та АТФ, а також калій 1 кг/га для стабілізації осмотичного балансу та роботи клітинних мембран.
Дотримання цих спеціалізованих схем дозволяє рослині ефективно підготуватися до стресових умов, мінімізувати пошкодження клітин, зберегти фотосинтетичну активність і забезпечити швидке відновлення росту після дії несприятливих факторів, що є ключем до підтримки високої продуктивності культури.
Таблиця 9
Схеми під конкретний стрес
| № | Тип стресу / Ситуація | Час застосування | Найефективніші речовини / норма | Примітка / ефект |
| 1 | Посуха, високі температури, засоленість, підвищена кислотність ґрунту | 3–5 днів до стресу | Калій — 2 кг/га, Амінокислоти — 1 л/га, Гліцин-бетаїн — 0,5 кг/га | Підвищує стійкість, зменшує втрати фотосинтезу та енергії |
| 2 | Гербіцидний стрес | 24–48 год після обробки | Амінокислоти — 1,5 л/га, Магній — 1 кг/га, Фосфор — 0,5 кг/га | Захист хлоропластів, зниження ROS, відновлення росту |
| 3 | Холод | 1–2 дні до стресу | Амінокислоти — 1 л/га, Фосфор — 1 кг/га, Калій — 1 кг/га | Підвищує морозостійкість та відновлення росту |
| 4 | Найефективніші речовини (рейтинг) | — | 1. Амінокислоти — дуже висока, 2. Магній — дуже висока, 3. Калій — дуже висока, 4. Гліцин-бетаїн — дуже висока, 5. Кремній — висока, 6. Фосфор — висока | Науково-практичне обґрунтування |
| 5 | Універсальна антистресова схема (Антистрес №1) | Для 1 га | Амінокислоти — 1,5 л, MgSO₄ — 1 кг, Калій — 1 кг | Зниження втрат врожаю на 20–50 % |
| 6 | Ключовий елемент антистресу | Постійно | Магній | Контролює фотосинтез, енергію та ROS |
| 7 | Професійна схема (світовий стандарт) | — | Амінокислоти + Mg + K | Використовується у ЄС, США, Бразилії |
| 8 | Практичний результат (приріст врожайності) | — | — | Пшениця: 0,3–0,8 т/га, Кукурудза: 0,5–1,5 т/га, Соняшник: 0,2–0,6 т/га |

Мал. 4. Метаболічний зсув при стресі.
Найважливішою практичною схемою універсального антистресового захисту рослин є застосування бакової суміші препаратів «Антистрес №1», яка забезпечує комплексну підтримку метаболізму, фотосинтезу та енергетичного балансу культури. На один гектар рекомендується внесення амінокислот у нормі 1,5 л для забезпечення рослини готовими субстратами для синтезу білків і ферментів, MgSO₄ 1 кг для відновлення роботи хлоропластів і синтезу АТФ, а також калію 1 кг для підтримки водного балансу, роботи продихів і стабілізації клітинних мембран. Така комплексна обробка дозволяє знизити втрати врожаю у періоди стресу на 20–50 %, що робить її універсальним рішенням для широкого спектру культур і умов вирощування.
Критично важливим компонентом цієї схеми є магній, який виступає ключовим елементом антистресової підтримки рослини. Він контролює роботу фотосинтезу, забезпечує синтез і накопичення енергії у вигляді АТФ та зменшує рівень реактивних форм кисню, що дозволяє клітинам ефективно використовувати наявні ресурси і мінімізувати оксидативні пошкодження.
Найефективніша схема в світі, яка відповідає професійному стандарту і застосовується в країнах ЄС, США та Бразилії, базується на простій, але дієвій формулі: амінокислоти + магній + калій. Вона забезпечує універсальну підтримку рослини у будь-яких стресових умовах і дозволяє досягти відчутного підвищення врожайності. Практичний результат застосування цієї схеми демонструє приріст продуктивності таких культур: пшениця — 0,3–0,8 т/га, кукурудза — 0,5–1,5 т/га, соняшник — 0,2–0,6 т/га. Таким чином, універсальна антистресова схема забезпечує стабільність росту, підвищує стійкість до стресових факторів і створює основу для реалізації максимального продуктивного потенціалу культур навіть у складних агрокліматичних умовах.
Реальні можливості традиційних антистресових препаратів і їх обмеження у регуляції стресостійкості рослин.
Стресова відповідь рослин контролюється фундаментальними сигнальними системами клітини, центральне місце серед яких займають енергетичні сенсори SnRK1 (SNF1-related kinase 1) і TOR (Target of Rapamycin). Ці системи визначають, чи клітина буде рости, чи переходити у режим виживання. Їхня активність регулюється не зовнішніми антистресовими препаратами як такими, а внутрішнім енергетичним (рівнем цукрів), метаболічним і гормональним станом клітини.
Останніми роками застосування антистресових препаратів у рослинництві набуло широкого поширення як інструмент підвищення продуктивності в умовах абіотичних і біотичних стресів. До цієї групи продуктів найчастіше відносять амінокислоти, екстракти морських водоростей, гумінові та фульвові кислоти, а також різні біостимулятори природного походження. Разом з тим їхня реальна роль у регуляції стресостійкості часто перебільшується, особливо у контексті фундаментальних механізмів клітинного контролю енергетичного метаболізму.
Центральними регуляторами клітинної відповіді на стрес є висококонсервативні сигнальні системи SnRK1 (SNF1-related kinase 1) і TOR (Target of Rapamycin), які функціонують як сенсори енергетичного статусу клітини. SnRK1 активується в умовах енергетичного дефіциту і переводить клітину у режим виживання, пригнічуючи біосинтетичні процеси і ріст. У протилежність цьому, TOR активується при достатньому енергетичному забезпеченні і стимулює синтез білка, ріст клітин і формування врожаю. Активність цих систем визначається насамперед внутрішніми метаболічними сигналами, такими як рівень цукрів, співвідношення АТФ/АМФ і гормональний баланс, а не екзогенними антистресовими препаратами.
Більшість комерційних антистресових продуктів на ринку не здатні безпосередньо регулювати активність SnRK1 або TOR. Їхній ефект реалізується переважно через непрямі механізми, зокрема стабілізацію клітинних мембран, часткове зниження оксидативного пошкодження, підтримання осмотичного балансу і прискорення відновлення метаболізму після стресу. Наприклад, амінокислоти можуть використовуватися як додаткове джерело азоту і субстрати для синтезу білків, екстракти водоростей містять низькі концентрації природних фітогормонів, а гумінові речовини можуть покращувати поглинання елементів живлення. Однак ці ефекти не означають прямого контролю над центральними енергетичними перемикачами клітини.
Фундаментальними факторами, що визначають стресостійкість і продуктивність, залишаються процеси фотосинтезу, транспорт і розподіл асимілятів у системі SOURCE (донори) – SINK (акцептори), а також гормональна регуляція росту. Саме рівень доступних вуглеводів, особливо сахарози і сигнальних цукрових метаболітів, визначає, чи буде активований шлях росту через TOR або стресова програма через SnRK1. Таким чином, ключовим обмежуючим фактором врожаю при стресі є не відсутність антистресових препаратів, а порушення енергетичного балансу і транспорту асимілятів.
Отже, антистресові препарати слід розглядати як допоміжний інструмент, який може частково пом’якшити наслідки стресу і прискорити відновлення рослин, але не як засіб прямого управління фундаментальними механізмами клітинної регуляції. Перебільшення їх ролі може призводити до неправильного розуміння фізіологічних основ формування врожаю. Ефективні стратегії підвищення стресостійкості повинні бути спрямовані насамперед на підтримання фотосинтетичної активності, оптимізацію мінерального живлення і збереження ефективного SOURCE–SINK балансу, що забезпечує стабільне енергетичне забезпечення рослини.
Разом з тим існує обмежена кількість молекул, які дійсно можуть регулювати ключові стресові сигнальні системи. Регуляція енергетичного стану клітини здійснюється внутрішніми сигнальними молекулами, такими як цукри і фітогормони, які контролюють активність сигнальних шляхів TOR і SnRK1. Комерційні антистресові препарати, включаючи амінокислоти, екстракти водоростей і гумінові речовини, не здатні безпосередньо керувати цими системами, але можуть частково зменшувати пошкодження клітин і прискорювати відновлення після стресу.
Таблиця 10
Центральні регулятори енергетичного стану та стресових програм у рослин
| Група молекул | Молекула | Основна функція | Вплив на TOR | Вплив на SnRK1 | Фізіологічний ефект |
| Цукрові сигнали | Trehalose-6-phosphate (T6P) | Центральний сенсор енергетичного статусу | Активує | Інгібує | Перемикання між ростом і виживанням; підтримка росту навіть при стресі |
| Глюкоза | Сигнальна молекула енергетичної доступності | Активує | Пригнічує | Стимуляція синтезу білка, підтримка росту | |
| Сахароза | Системний сигнал SOURCE–SINK | Опосередкований вплив | Опосередкований вплив | Регуляція генів стресостійкості; контроль балансу джерело–стік | |
| Фітогормони | Абсцизова кислота (ABA) | Головний гормон стресу | Пригнічує (через стресові шляхи) | Активує (опосередковано) | Перехід у режим виживання; закриття продихів; активація генів стресу |
| Цитокініни | Антагоністи стресового старіння | Активують | Опосередковано пригнічують | Підтримка TOR-залежного росту; активація sink-органів | |
| Ауксини | Регуляція перерозподілу ресурсів | Непрямий вплив | Непрямий вплив | Розвиток кореневої системи; оптимізація ресурсів | |
| Етилен | Регулятор старіння і пошкодження | Пригнічує ріст | Активує стресові шляхи | Старіння; відповідь на ушкодження | |
| Енергетичні метаболіти | АТФ | Прямий показник енергетичного рівня | Активує (високий рівень) | Пригнічує (високий рівень) | Високий АТФ — ріст; низький — запуск режиму економії |
| AMP | Сигнал енергетичного дефіциту | Пригнічує | Активує | Перехід у катаболічний режим | |
| Сигнальні окисні молекули | ROS | Редокс-сигнали стресу | Непрямий вплив | Непрямий вплив | Регуляція генів стресової відповіді |
| Молекули без прямого контролю TOR/SnRK1 | Амінокислоти | Метаболіти / будівельні блоки | Не керують безпосередньо | Не керують безпосередньо | Можуть зменшувати пошкодження, але не запускають центральні енергетичні програми |
| Екстракти водоростей | Біостимулятори | Непрямий ефект | Непрямий ефект | Стабілізація мембран, антистресовий ефект | |
| Гумінові речовини | Структурні стимулятори | Непрямий ефект | Непрямий ефект | Поліпшення поглинання, відновлення | |
| Фульвові кислоти | Комплексоутворювачі | Непрямий ефект | Непрямий ефект | Покращення транспорту елементів |
SOURCE–SINK balance як центральний механізм стресостійкості і формування врожаю.
Формування врожаю сільськогосподарських культур визначається не лише інтенсивністю фотосинтезу, але насамперед ефективністю транспорту і розподілу асимілятів між органами-донорами (SOURCE) і органами-акцепторами (SINK). Концепція SOURCE–SINK балансу є фундаментальною основою продуктивності рослин і визначає їхню здатність підтримувати ріст і формування репродуктивних органів, особливо в умовах стресу.
SOURCE-органами є фотосинтезуючі листки, в яких відбувається фіксація CO₂ і синтез сахарози як основної транспортної форми вуглецю. SINK-органи включають точки росту, кореневу систему, молоді листки і, найважливіше, репродуктивні структури, зокрема зерно, яке є кінцевим акцептором асимілятів. Врожайність безпосередньо залежить від здатності SINK-органів ефективно імпортувати і використовувати сахарозу.
Центральним параметром, що визначає ефективність цього процесу, є так звана сила акцептора (sink strength), яка характеризує здатність тканини імпортувати і метаболізувати асиміляти. Ця властивість визначається активністю транспортерів сахарози, зокрема білків родин SUT і SWEET, а також ферменту cell wall invertase, який каталізує необоротний гідроліз сахарози на глюкозу і фруктозу в апопласті. Розщеплення сахарози підтримує концентраційний градієнт, необхідний для безперервного транспорту асимілятів у клітини акцепторних органів і забезпечує накопичення запасних речовин у зерні.
В умовах стресу, зокрема посухи, високих температур або дефіциту мінерального живлення, одним із перших порушується саме SOURCE–SINK баланс. Це проявляється у зниженні фотосинтетичної активності SOURCE-органів, пригніченні транспорту сахарози через флоему і зниженні активності ферментів, що визначають sink strength. У результаті навіть за наявності певного рівня фотосинтезу ефективність транспорту асимілятів до репродуктивних органів різко знижується, що призводить до порушення наливу зерна і втрат врожаю.
Важливою особливістю є те, що зниження врожайності при стресі часто пов’язане не стільки зі зменшенням фотосинтезу як таким, скільки з порушенням використання асимілятів акцепторними органами. Пригнічення активності cell wall invertase і транспортерів сахарози призводить до зменшення sink strength, що викликає накопичення вуглеводів у листках і одночасний дефіцит асимілятів у зерні. Таким чином, ключовим обмежуючим фактором продуктивності стає не синтез, а ефективний розподіл продуктів фотосинтезу.
SOURCE–SINK баланс також тісно пов’язаний із центральними системами клітинної регуляції енергетичного статусу, зокрема сигнальними шляхами TOR і SnRK1. При достатньому надходженні асимілятів активується TOR-залежний шлях, який стимулює синтез білка, клітинний поділ і ріст акцепторних органів. У протилежність цьому, при дефіциті асимілятів активується SnRK1, який пригнічує анаболічні процеси і переводить клітину у режим збереження енергії. Таким чином, ефективність SOURCE–SINK взаємодії безпосередньо визначає, чи буде рослина продовжувати формування врожаю або переходити у стан стресового пригнічення росту.
Отже, підтримання ефективного SOURCE–SINK балансу є центральною умовою стресостійкості і стабільної продуктивності рослин. Збереження фотосинтетичної активності SOURCE-органів, ефективного транспорту сахарози і високої сили акцепторних органів забезпечує безперервне постачання енергії і пластичних речовин до репродуктивних структур, що дозволяє мінімізувати негативний вплив стресових факторів на врожайність. У цьому контексті SOURCE–SINK система виступає ключовим інтегруючим механізмом, який поєднує процеси фотосинтезу, метаболізму і росту в єдину функціональну мережу, що визначає кінцеву продуктивність рослин.
Головний механізм антистресової стійкості рослин полягає у збереженні балансу SOURCE–SINK. Усі ефективні антистресові технології працюють саме через підтримання цього співвідношення, що включає збереження фотосинтетичної активності (SOURCE), стабільність транспорту асимілянтів та підтримання розвитку зерна або інших органів-акцепторів (SINK).
Практичне значення цього принципу очевидне: будь-який антистресовий препарат має або зберігати функціональність SOURCE, або підсилювати SINK, або оптимізувати транспорт між ними. Якщо ж баланс SOURCE–SINK не підтримується, зростання врожайності неможливе, незалежно від сили стресового чинника чи застосованих засобів.
Ключовий фундаментальний висновок полягає в тому, що головною причиною втрати врожаю за умов стресу є не сам стресовий фактор, а викликане ним порушення SOURCE–SINK балансу. Отже, керування цим балансом фактично означає керування врожайністю.
Мал. 5. Антистрес = не амінокислоти, а збереження SOURCE–SINK. Антистрес працює лише якщо збережено потік вуглецю SOURCE → SINK. Будь-який антистресовий препарат повинен зберігати SOURCE або підсилювати SINK або покращувати транспорт. Якщо цього не відбувається — врожай не зростає. Хто керує SOURCE–SINK balance — той керує врожаєм.
Таблиця 11
SOURCE–SINK balance як центральний механізм стресостійкості та формування врожаю
| Рівень регуляції | Компонент | Локалізація | Функція в системі SOURCE–SINK | Вплив стресу | Наслідок для врожаю |
| SOURCE | Фотосинтез (CO₂ → сахароза) | Листки | Синтез транспортної форми вуглецю (сахарози) | ↓ фотосинтезу | ↓ загального потоку асимілятів |
| Транспорт | Флоемний транспорт | Провідна система | Переміщення сахарози від SOURCE до SINK | Порушення провідності | ↓ надходження асимілятів у зерно |
| Транспортери | SUT (Sucrose Transporters) | Мембрани клітин флоеми та SINK | Активний імпорт сахарози | ↓ експресії/активності | Ослаблення sink strength |
| SWEET transporters | Мембрани клітин SOURCE/SINK | Пасивний експорт та міжклітинний транспорт сахарози | Дисбаланс експорту | Порушення градієнта транспорту | |
| Ферментативний вузол | Cell wall invertase | Апопласт SINK-органів | Гідроліз сахарози → глюкоза + фруктоза; підтримання концентраційного градієнта | ↓ активності | ↓ сили акцептора, порушення наливу зерна |
| Метаболічний вузол | Sink strength | Зерно, меристеми | Здатність імпортувати та утилізувати асиміляти | Ослаблення через ↓ транспортерів/інвертази | Прямі втрати врожаю |
| Енергетична інтеграція | TOR | Клітини SINK | Активація росту при достатньому потоці асимілятів | Інгібування при дефіциті | Пригнічення росту |
| SnRK1 | Універсальний сенсор дефіциту | Перехід у режим економії енергії | Гіперактивація | Блокування анаболізму | |
| Системний результат | SOURCE–SINK balance | Уся рослина | Інтеграція синтезу, транспорту і використання | Порушення балансу | Основна причина втрати врожаю |
Молекулярні механізми регуляції SOURCE–SINK балансу.
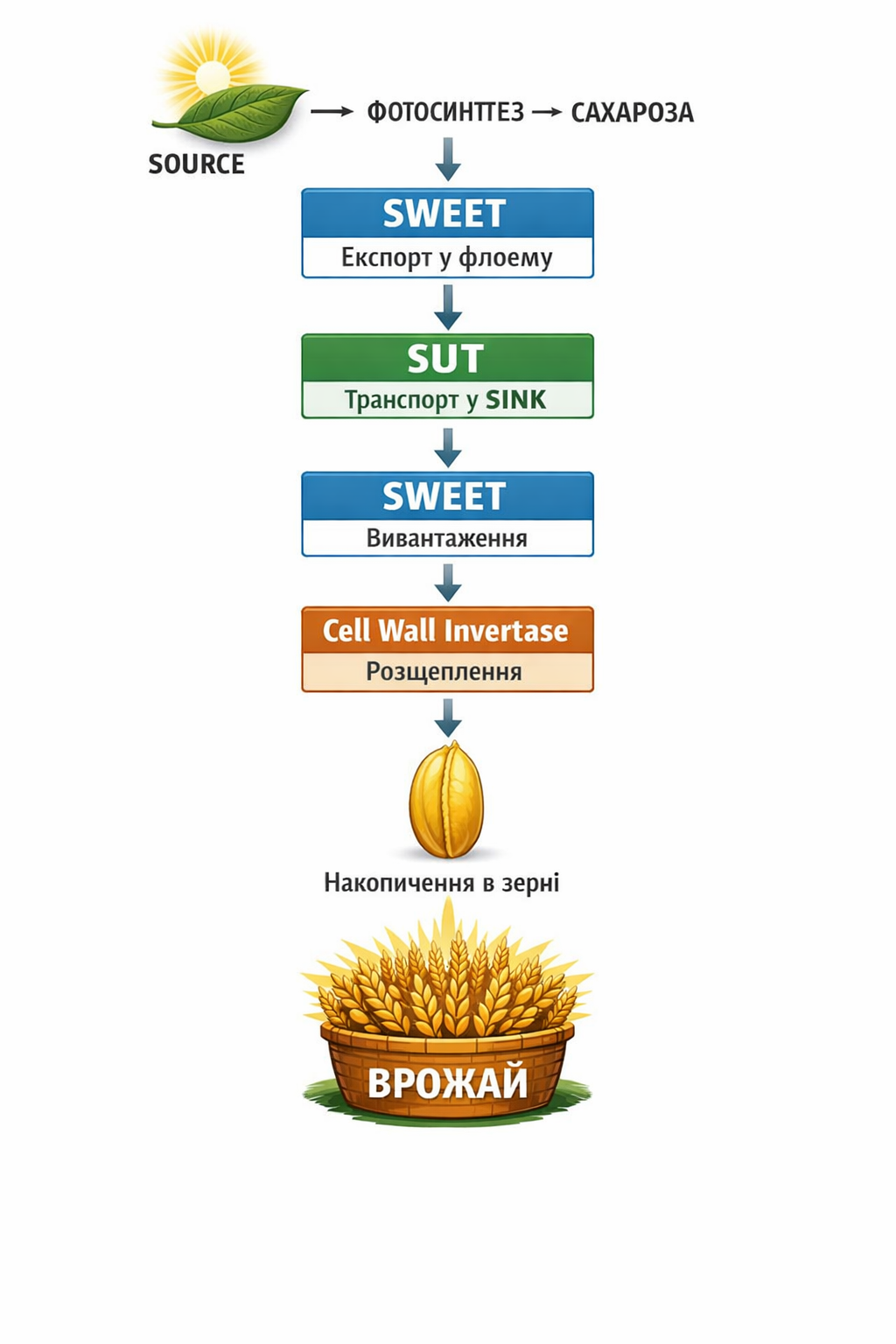
Врожай — це функція транспорту сахарози, а не тільки фотосинтезу. Молекулярні механізми регуляції SOURCE–SINK балансу визначають ефективність формування врожаю через контроль завантаження, транспорту, реабсорбції та розвантаження сахарози між органами-донорами (SOURCE) і органами-акцепторами (SINK). Центральним процесом є переміщення сахарози як основної транспортної форми вуглецю, що забезпечується спеціалізованими мембранними транспортерами та ферментами, які регулюють її рух і необоротне використання в тканинах.
Ключову роль відіграють транспортери SUT (Sucrose–H⁺ symporters) — активні білки плазматичної мембрани, які здійснюють енергозалежний транспорт сахарози разом із протонами. Їхня робота базується на протонному градієнті (H⁺ gradient), створеному H⁺-АТФазами. Механістично процес можна представити як: H⁺ + sucrose → транспорт у клітину. У листках SUT забезпечують завантаження сахарози у флоему (phloem loading), визначаючи швидкість експорту вуглецю з SOURCE. Під час флоемного транспорту вони також здійснюють реабсорбцію сахарози, запобігаючи її втратам і підтримуючи ефективність потоку. У SINK-органах — зерні, коренях і меристемах — SUT забезпечують активний імпорт сахарози з флоеми до клітин-акцепторів. Висока експресія SUT корелює зі збільшенням надходження вуглецю до зерна та зростанням його маси, тоді як посуха й температурний стрес знижують їх експресію, що призводить до метаболічного «голодування» зерна навіть за відносно збереженого фотосинтезу.
Другим критично важливим компонентом є транспортери SWEET, які забезпечують пасивний транспорт сахарози за градієнтом концентрації (facilitated diffusion). Вони здійснюють експорт сахарози з клітин SOURCE в апопласт і відіграють центральну роль у phloem unloading — вивантаженні сахарози в SINK-органах. У зерні, зокрема в ендоспермі, SWEET транспортують сахарозу з флоеми до клітин накопичення, визначаючи швидкість наповнення зернівки. Стресові фактори знижують їх активність, що безпосередньо обмежує потік вуглецю до репродуктивних органів і зменшує швидкість наливу.
Третій компонент — інвертази (invertases), ферменти, що каталізують гідроліз сахарози за реакцією: Sucrose → Glucose + Fructose. Цей процес є принципово важливим, оскільки гексози не можуть повернутися у флоему, що створює концентраційний градієнт і підтримує безперервний імпорт нової сахарози. Таким чином формується так званий механізм метаболічної «пастки» (metabolic trap mechanism), який лежить в основі сили акцептора (sink strength). Існують три основні типи інвертаз: цитозольні, вакуолярні та апопластичні (cell wall invertase).
Найважливішою для формування врожаю є cell wall invertase (CWIN), локалізована в апопласті між флоемою та клітинами зерна. CWIN – це головний фермент врожаю. Вона здійснює необоротне розщеплення сахарози до гексоз у зоні її надходження до SINK-органа, підтримуючи постійний градієнт імпорту. Саме CWIN формує метаболічну основу sink strength, визначає швидкість наливу зерна, його розмір і кінцеву врожайність. Експериментальні дані свідчать, що зниження активності CWIN може зменшувати врожай на 30–60 %, а в умовах посухи її пригнічення пов’язане з абортацією зерна. Крім того, гексози, утворені внаслідок дії CWIN, є субстратами для синтезу крохмалю в ендоспермі, що безпосередньо визначає масу зернівки.
Функціонування цієї транспортно-ферментативної системи інтегрується з енергетичними сигнальними шляхами TOR і SnRK1. За достатнього потоку асимілятів активується TOR-залежна регуляція, що стимулює синтез білка, клітинний поділ і крохмалеутворення в зерні. При дефіциті вуглецю активується SnRK1, який пригнічує анаболічні процеси та переводить клітину в режим енергозбереження, знижуючи sink strength навіть за наявності часткового транспорту сахарози.
Інтегрований механізм формування врожаю можна представити як послідовність: фотосинтез у SOURCE → синтез сахарози → експорт через SWEET → завантаження і підтримка потоку за участю SUT → вивантаження через SWEET у SINK → необоротний гідроліз сахарози cell wall invertase → утворення гексоз → синтез крохмалю → накопичення в зерні → врожай.
За умов стресу пригнічується експресія SUT і SWEET, знижується активність інвертаз, особливо CWIN, активується SnRK1 та інгібується TOR. У результаті порушується імпорт і метаболічна фіксація вуглецю, що призводить до різкого зменшення врожайності навіть без повної зупинки фотосинтезу.
Отже, сила акцептора визначається не стільки рівнем фотосинтезу, скільки інтегрованою активністю транспортерів SUT і SWEET, ферменту cell wall invertase та їх координацією з енергетичними сигнальними системами. Врожай є функцією ефективного транспорту, метаболічної «фіксації» і перерозподілу вуглецю, а не лише інтенсивності фотосинтезу.
Таблиця 12
Молекулярні механізми формування сили акцептора (sink strength)
| Молекулярний компонент | Тип білка / ферменту | Локалізація | Основна функція | Роль у формуванні sink strength |
| SUT (Sucrose Transporters) | Мембранні транспортери сахарози | Плазматична мембрана клітин флоеми та SINK | Активний імпорт сахарози у клітину | Забезпечують накопичення сахарози в акцепторній тканині |
| SWEET transporters | Пасивні транспортери (фасилітатори) | Плазматична мембрана клітин SOURCE і SINK | Експорт/перерозподіл сахарози між клітинами | Формують концентраційний градієнт транспорту |
| Invertases (загальна група) | Гідролітичні ферменти | Вакуоля, цитозоль, апопласт | Розщеплення сахарози на глюкозу і фруктозу | Підтримують використання асимілятів у тканині |
| Cell wall invertase (CWIN) | Апопластична інвертаза | Клітинна стінка (апопласт) SINK-органів | Необоротний гідроліз сахарози в апопласті | Підтримує градієнт імпорту і є ключовим регулятором сили акцептора |
Чому більшість антистресових препаратів мають обмежену ефективність: погляд через призму SOURCE–SINK фізіології.
У сучасному рослинництві широко використовуються різноманітні антистресові препарати, включаючи амінокислоти, екстракти морських водоростей, гумінові і фульвові кислоти, а також інші біостимулятори. Їх застосування часто позиціонується як ефективний інструмент управління фізіологічним станом рослин і підвищення врожайності в умовах стресу. Проте з позиції фізіології SOURCE–SINK взаємодії ефективність більшості таких препаратів має об’єктивні обмеження.
Основною причиною зниження врожаю при стресі є порушення балансу між процесами утворення асимілятів у SOURCE-органах і їх використанням у SINK-органах. Стресові фактори, такі як посуха, високі температури або дефіцит елементів живлення, насамперед знижують фотосинтетичну активність листків, порушують транспорт сахарози через флоему і пригнічують метаболічну активність репродуктивних органів. У результаті зменшується надходження асимілятів до зерна, що безпосередньо обмежує його ріст і масу.
Більшість антистресових препаратів не здатні безпосередньо впливати на ці фундаментальні процеси. Наприклад, екзогенні амінокислоти можуть використовуватися клітиною як додаткові метаболічні субстрати, а гумінові речовини можуть частково покращувати поглинання мінеральних елементів, однак вони не можуть компенсувати дефіцит асимілятів, що виникає внаслідок пригнічення фотосинтезу. Аналогічно, біостимулятори на основі водоростей можуть містити низькі концентрації природних регуляторів росту, але їх вплив є допоміжним і не здатний повністю відновити транспорт і розподіл вуглецю при значному стресі.
Ключовим обмежуючим фактором є те, що формування врожаю визначається не наявністю окремих метаболітів, а інтегрованою функцією всієї SOURCE–SINK системи. Якщо фотосинтетичний апарат пошкоджений або транспорт асимілятів порушений, додаткове внесення амінокислот або інших органічних сполук не може повністю замінити втрачений потік вуглецю. В таких умовах головним завданням є відновлення функціональної активності SOURCE і забезпечення ефективного транспорту асимілятів до SINK, а не лише підтримка окремих метаболічних процесів.
Разом з тим це не означає, що антистресові препарати є повністю неефективними. Їх позитивний вплив може проявлятися у зменшенні ступеня клітинного пошкодження, стабілізації мембран, підтриманні ферментативної активності і прискоренні відновлення після припинення дії стресу. У сприятливих або помірно стресових умовах ці ефекти можуть сприяти кращому збереженню SOURCE–SINK функціонування. Однак їх дія є опосередкованою і не може розглядатися як прямий механізм управління формуванням врожаю.
Таким чином, з фізіологічної точки зору антистресові препарати не є засобом прямого контролю продуктивності, а лише допоміжним фактором, який може підвищити стабільність функціонування рослинної системи. Основними визначальними факторами врожаю залишаються інтенсивність фотосинтезу, ефективність транспорту асимілятів і активність акцепторних органів. Саме ці процеси формують фізіологічну основу продуктивності, тоді як антистресові препарати можуть лише частково підтримувати їх функціонування, але не замінювати.
Практичні агрономічні стратегії управління SOURCE–SINK балансом для підвищення врожаю.
Практичні агрономічні стратегії управління SOURCE–SINK балансом є одним із ключових інструментів підвищення врожайності та стабільності продуктивності рослин, особливо в умовах абіотичних і біотичних стресів. Формування врожаю є результатом узгодженої взаємодії трьох фундаментальних процесів: фотосинтезу, транспорту асимілятів і їх використання акцепторними органами. Тому ефективна агрономічна система повинна бути спрямована не на окреме стимулювання одного з цих процесів, а на підтримання функціональної рівноваги між SOURCE- та SINK-системами протягом критичних фаз розвитку культури.
Збереження фотосинтетичної активності SOURCE-органів.
Першочерговим завданням є збереження фотосинтетичної активності SOURCE-органів, насамперед листкового апарату, який виступає основним джерелом асимілятів. Будь-яке зниження фотосинтезу безпосередньо обмежує енергетичне забезпечення формування врожаю. Практично це досягається через оптимізацію азотного живлення, оскільки азот є ключовим компонентом хлорофілу та ферменту Rubisco. Його дефіцит прискорює старіння листків і зменшує інтенсивність фотосинтезу, тоді як оптимальне забезпечення дозволяє продовжити активність SOURCE-системи.
Не менш важливим є забезпечення рослин магнієм — центральним атомом молекули хлорофілу, який бере участь у фотохімічних реакціях. Захист листкового апарату від хвороб також має критичне значення, адже навіть часткове ураження істотно знижує загальну продукцію асимілятів. Підтримання водного балансу є ще одним необхідним компонентом: водний дефіцит викликає закриття продихів, обмежує надходження CO₂ і різко знижує фотосинтетичну продуктивність.
Гормональний контроль SOURCE–SINK.
Гормональний контроль SOURCE–SINK балансу є ключовим регуляторним механізмом формування врожаю та адаптації до стресу. Баланс між органами-донорами і органами-акцепторами координується системою фітогормонів, які визначають як інтенсивність транспорту асимілятів, так і силу акцептора. Цитокініни стимулюють sink strength, активуючи клітинний поділ і метаболічну активність у репродуктивних органах, а також затримують старіння листків, підтримуючи функціональність SOURCE. Ауксини регулюють полярний транспорт і перерозподіл асимілятів, впливаючи на напрямок їхнього руху до активно ростучих тканин. Абсцизова кислота (ABA) за умов стресу пригнічує активність SINK-органів, обмежуючи ріст і сприяючи переходу рослини в режим збереження ресурсів. Гібереліни, навпаки, стимулюють ріст зерна та інших акцепторних структур, посилюючи використання імпортованих асимілятів. Таким чином, гормональна система інтегрує сигнали розвитку і стресу, визначаючи ефективність SOURCE–SINK взаємодії та кінцеву продуктивність рослини.
Забезпечення ефективного транспорту асимілятів.
Другим критичним елементом є забезпечення ефективного транспорту асимілятів, передусім сахарози, через флоему від SOURCE до SINK. У цьому процесі ключову роль відіграє калій. Оптимальне калійне живлення є фундаментальною умовою стабільного транспорту, оскільки калій підтримує тургор клітин флоеми, забезпечує функціонування транспортерів сахарози та регулює осмотичні процеси. Дефіцит калію призводить до накопичення цукрів у листках та їх недостатнього надходження до зерна, що безпосередньо обмежує врожайність.
Підтримання сили акцептора (sink strength).
Навіть за достатнього фотосинтезу врожай може бути лімітований силою акцептора (sink strength), тобто здатністю репродуктивних органів ефективно імпортувати та використовувати асиміляти. Особливе значення має забезпечення рослин бором, який відіграє важливу роль у функціонуванні репродуктивних структур і транспорті вуглеводів. Його дефіцит призводить до погіршення наливу зерна та абортації генеративних органів. Водночас азотне живлення повинно бути збалансованим, оскільки як дефіцит, так і надлишок азоту порушують SOURCE–SINK рівновагу. Надлишок азоту стимулює надмірний вегетативний ріст і знижує пріоритет репродуктивних органів як основних SINK-структур.
Захист критичних фаз формування врожаю.
Особливої уваги потребує захист критичних фаз формування врожаю — колосіння, цвітіння та наливу зерна. Саме в цей період визначаються кількість зерен і їх маса, а будь-який стрес має максимальний негативний вплив на продуктивність. Порушення SOURCE–SINK взаємодії в ці фази практично не компенсується в подальшому.
Управління листковою тривалістю життя (stay-green ефект).
Важливим напрямом є управління тривалістю життя листків, так званий stay-green ефект. Чим довше листковий апарат залишається функціонально активним після цвітіння, тим більший обсяг асимілятів може бути спрямований до зерна. Подовження активності SOURCE безпосередньо підвищує потенціал наливу і стабільність врожаю.
Мінімізація факторів, що порушують SOURCE–SINK баланс.
Необхідно також мінімізувати фактори, що порушують SOURCE–SINK баланс, зокрема посуху, високі температури, дефіцит елементів живлення та хвороби. Їх негативний вплив реалізується передусім через порушення транспорту асимілятів і зниження активності акцепторних органів.
Практичний фізіологічний принцип управління врожаєм.
З практичної фізіологічної точки зору врожай визначається трьома взаємопов’язаними процесами: інтенсивністю фотосинтезу (SOURCE), ефективністю транспорту асимілятів і активністю та силою акцепторних органів (SINK). Порушення будь-якого з цих компонентів неминуче призводить до зниження продуктивності.
Отже, ефективні агрономічні стратегії підвищення врожаю повинні бути спрямовані не лише на стимулювання фотосинтезу, а насамперед на підтримання збалансованої взаємодії між SOURCE і SINK системами. Оптимізація мінерального живлення, захист листкового апарату та забезпечення повноцінного функціонування репродуктивних органів дозволяють зберігати ефективний транспорт і використання асимілятів навіть в умовах стресу. Саме управління SOURCE–SINK балансом є одним із найбільш фундаментальних і дієвих механізмів стабілізації та підвищення врожайності.
Таблиця 13
Практичні агрономічні стратегії управління SOURCE–SINK балансом
| № | Напрям управління | Конкретні заходи | Фізіологічний механізм | Ризики при порушенні |
| 1 | Збереження фотосинтетичної активності (SOURCE) | Оптимізація азотного живлення | Підтримка синтезу хлорофілу та Rubisco, продовження активності листків | Передчасне старіння, зниження фотосинтезу |
| Забезпечення магнієм | Участь Mg у структурі хлорофілу та фотохімічних реакціях | Зниження ефективності фотосинтезу | ||
| Захист від хвороб | Збереження площі активного листкового апарату | Втрата продукції асимілятів | ||
| Підтримання водного балансу | Запобігання закриттю продихів, підтримка надходження CO₂ | Різке зниження фотосинтезу | ||
| 2 | Ефективний транспорт асимілятів | Оптимальне калійне живлення | Підтримка тургору флоеми, робота транспортерів сахарози, осмотичний транспорт | Накопичення цукрів у листках, поганий налив зерна |
| 3 | Підтримання сили акцептора (SINK strength) | Забезпечення бором | Регуляція транспорту вуглеводів, функціонування репродуктивних органів | Абортація квіток, слабкий налив зерна |
| Збалансоване азотне живлення | Узгодження вегетативного та репродуктивного росту | Надлишок N → надмірний вегетативний ріст, ослаблення SINK | ||
| 4 | Захист критичних фаз розвитку | Контроль стресів у фазах колосіння, цвітіння, наливу зерна | Збереження кількості зерен та їх маси | Максимальні втрати врожаю при стресі |
| 5 | Stay-green ефект | Подовження функціонування листків після цвітіння | Триваліший потік асимілятів у зерно | Передчасна втрата SOURCE-функції |
| 6 | Мінімізація стресових факторів | Контроль посухи, температур, живлення, хвороб | Підтримка транспорту асимілятів і активності SINK | Порушення SOURCE–SINK взаємодії |
Таблиця 14
Практичний фізіологічний принцип управління врожаєм
| Компонент | Роль у формуванні врожаю | Наслідок порушення |
| SOURCE (інтенсивність фотосинтезу) | Формування асимілятів | Енергетичний дефіцит |
| Транспорт | Переміщення сахарози через флоему | Накопичення цукрів у листках |
| SINK (сила акцептора) | Використання асимілятів для формування зерна | Недостатній налив, зниження маси |
Мал. 6. Енергетична дестабілізація при абіотичному стресі.
Молекулярні механізми регуляції SOURCE–SINK балансу включають завантаження, транспорт і розвантаження сахарози між SOURCE- і SINK-органами, що визначає ефективність формування врожаю. Центральну роль у цьому процесі відіграють мембранні транспортери та ферменти: SUT (Sucrose Transporters, активний H⁺-симпорт сахарози), SWEET (пасивний facilitated diffusion), інвертази та Cell wall invertase (CWIN). SUT забезпечують завантаження сахарози у флоему та транспорт у SINK-тканини, використовуючи протонний градієнт, а SWEET відповідають за вивантаження сахарози в апопласт SINK-органів. Інвертази, особливо CWIN, розщеплюють сахарозу на глюкозу і фруктозу, що створює концентраційний градієнт і забезпечує необоротне накопичення асимілятів у зерні, визначаючи sink strength. Зниження активності цих білків під дією стресу призводить до зменшення наливу зерна та втрати врожаю навіть при відносно збереженому фотосинтезі.
Фізіологічний рівень інтегрує pressure-flow mechanism, коли тургорний градієнт між SOURCE і SINK рухає флоемний потік, а активність транспортерів та CWIN визначає швидкість і напрям потоку. Гормональна регуляція координує активність транспортерів: цитокініни підсилюють sink strength і затримують старіння листків, ауксини регулюють транспорт, ABA пригнічує SINK при стресі, а гібереліни стимулюють ріст зерна. При дефіциті асимілятів активується SnRK1, який пригнічує анаболічні процеси, тоді як TOR стимулює синтез білка та клітинний поділ, інтегруючи енергетичний та гормональний контроль.
Ефективність SOURCE–SINK балансу також залежить від C/N співвідношення, оскільки транспорт і використання азотних та вуглецевих асимілятів координуються для підтримки росту і розвитку репродуктивних органів. Крім того, високий рівень SINK органів забезпечує feedback inhibition фотосинтезу в SOURCE, формуючи динамічну систему регуляції, де фотосинтетична активність і транспорт асимілятів взаємно підсилюють або гальмують один одного.
Sink strength формально визначається як добуток розміру акцепторного органа на його здатність активно імпортувати і метаболізувати асиміляти: Sink strength = sink size × sink activity. Саме активність SUT, SWEET та CWIN визначає кінцевий врожай, а порушення цього балансу під стресом стає головною причиною зниження продуктивності. Таким чином, підтримання фотосинтезу в SOURCE, ефективного транспорту через флоему, гормональної координації та стабільної активності ключових ферментів у SINK забезпечує максимальний урожай і є основним механізмом антистресової стійкості рослин.
Цитокініни як ключові регулятори SOURCE–SINK і sink strength.
Цитокініни — найважливіший клас фітогормонів, що контролюють баланс SOURCE–SINK і визначають силу SINK. Вони підвищують експресію генів SUT, активність cell wall invertase, стимулюють налив зерна і загальну продуктивність. Науково доведено, що цитокініни здатні підвищувати активність інвертаз на 30–80 %. Практичні препарати включають 6-BAP (6-бензиламінопурин), Zeatin, Kinetin та екстракти морських водоростей (Ascophyllum nodosum), які забезпечують більший розмір зерна, швидший налив і підвищення врожаю.
6-BAP — синтетичний цитокінін, що імітує природні молекули, стимулюючи клітинне ділення в меристемах, регулюючи баланс SOURCE–SINK і уповільнюючи листяне старіння. Використовується для стимуляції бруньок, інтенсивного цвітіння, формування плодів і в насінництві in vitro. Ефективний у низьких концентраціях (0,01–0,1 мг/л in vitro; 10–50 мг/л у полі), надлишок може викликати патологічне розгалуження та пригнічення кореневої системи.
Zeatin — природний цитокінін, особливо з кукурудзи, стимулює клітинний поділ і диференціацію, підвищує фотосинтетичну активність, зберігає хлорофіл, затримує старіння листя і розвиток бруньок. Застосовується для утворення пагонів, кореневих систем, підвищення стійкості до абіотичних стресів та продовження вегетації. Біологічно активний у концентраціях 0,01–10 мг/л, найефективніший у комбінації з іншими гормонами.
Kinetin — синтетичний цитокінін, що стимулює поділ клітин і диференціацію, уповільнює старіння листя та пагонів, покращує водний баланс і стійкість до стресів. Використовується як у лабораторних культурах, так і в польових умовах для стимуляції росту пагонів і листя, підвищення урожайності. Найефективніший у комбінації з 6-BAP або ауксинами; надлишок може викликати нерівномірний ріст.
Екстракти морських водоростей (Ascophyllum nodosum) — природний біостимулятор, що містить цитокініни, ауксини, абсцизову кислоту, полісахариди та мікроелементи. Вони підвищують стійкість рослин до стресів, покращують метаболізм, розвиток кореневої системи і фотосинтез. Використовуються для обприскування під час стресових умов, підвищують врожайність і якість плодів. М’які, безпечні, дозування залежить від культури (зазвичай 1–5 л/га) і можуть поєднуватися з іншими агрохімікатами.
Таким чином, цитокініни є ключовими регуляторами sink strength, забезпечуючи ефективний перенос асимілятів із SOURCE до SINK, оптимізуючи налив зерна та загальну продуктивність рослин.
Таблиця 15
Цитокініни як регулятори сили акцепторних органів і наливу зерна
| Препарат | Тип | Механізм дії | Основні ефекти | Форма випуску | Рекомендоване дозування | Особливості застосування |
| 6-BAP (6-бензиламінопурин) | Синтетичний цитокінін | Стимулює клітинний поділ у меристемах, регулює source–sink, зменшує старіння листя | Утворення додаткових пагонів, стимуляція цвітіння, підвищення зав’язування плодів | Порошок, розчин для обприскування | 10–50 мг/л (польові культури), 0,01–0,1 мг/л (in vitro) | Надлишок може пригнічувати кореневу систему та викликати нерівномірне розгалуження |
| Zeatin (Зеатин) | Природний цитокінін | Сприяє клітинному поділу та диференціації, підтримує фотосинтез, уповільнює сенесценцію | Стимуляція росту пагонів, підвищення стресостійкості, продовження вегетації | Водний або спиртовий розчин, порошок | 0,01–10 мг/л | Найефективніший у комбінації з іншими гормонами |
| Kinetin (Кінетин) | Синтетичний цитокінін | Стимулює клітинний поділ, уповільнює старіння, покращує водний баланс | Ріст пагонів та листя, підвищення врожайності | Порошок, водний розчин для обприскувань | 10–50 мг/л | Ефективний у комбінації з 6-BAP або ауксинами; надлишок може викликати нерівномірний ріст |
| Seaweed extracts (Ascophyllum nodosum) | Природний біостимулятор | Містить природні гормони, полісахариди, мікроелементи; підвищує стресостійкість | Стимуляція росту коренів та пагонів, підвищення врожайності і якості плодів, антистресовий ефект | Рідкі концентрати для розведення водою | 1–5 л/га | Безпечний, нетоксичний; можна поєднувати з іншими препаратами |
На основі 6-BAP, Zeatin та Kinetin створено численні комерційні препарати, які активно використовуються у сільському господарстві, тепличному виробництві та біотехнології рослин. Препарати на основі 6-BAP (6-бензиламінопурин) представлені такими відомими комерційними продуктами, як MaxCel® (Valent BioSciences), Promalin® (BASF, містить 6-BAP + GA₄₊₇), Perlan® та Accel®. Вони застосовуються для збільшення розміру плодів яблуні, проріджування груш, покращення якості ягід винограду та стимуляції росту овочевих культур. Типові норми використання становлять 20–100 ppm.
Zeatin, будучи дорогим природним гормоном, частіше входить до складу преміум-біостимуляторів та лабораторних середовищ, зокрема MS medium, а також екстрактів морських водоростей і середовищ для тканинної культури. Препарати на основі Kinetin застосовуються переважно як лабораторні реактиви та біостимулятори для стимуляції росту, уповільнення старіння (anti-senescence) і тканинної культури рослин.
Важливо зазначити, що більшість польових біостимуляторів містять цитокініни, наприклад, Kelpak (Ascophyllum nodosum), Megafol (морські водорості), Stimulate (містить kinetin) та Cytokin (синтетичний цитокінін). На практиці в полі рідко застосовують чистий 6-BAP через його високу ціну, нестабільність і складність використання; натомість ефективніше використовувати біостимулятори на основі морських водоростей, які дешевші, стабільніші та безпечніші.
Серед реальних препаратів, що застосовуються на польових культурах, виділяються Kelpak (natural cytokinin), Goëmar (seaweed cytokinin), Algaren (natural cytokinin) та Cytokin (synthetic). Всі ці засоби діють через ключовий механізм — посилення балансу SOURCE → SINK, тобто забезпечення ефективного перенесення асимілятів із листя до колоса або зерна.
На пшениці використання цитокінінових препаратів демонструє значний ефект: збільшення кількості зерен, маси зерна та загальної врожайності, зазвичай у межах +5–20 %. Найбільш практичним та економічно вигідним варіантом для польових культур є застосування екстрактів морських водоростей, оскільки вони поєднують стабільність, безпечність та ефективність при помірній вартості.
Таким чином, усі цитокінінові препарати працюють через фундаментальний механізм регуляції sink strength, забезпечуючи ефективний транспорт асимілятів від SOURCE до SINK і прямо впливаючи на формування врожаю.
Таблиця 16
Препаратів із цитокінінами для пшениці з
поясненням механізму дії на рівні SOURCE–SINK
| № | Препарат | Виробник | Активна речовина / Основа | Норма застосування | Фаза | Ефект на пшениці | Механізм дії (SOURCE → SINK) |
| 1 | Kelpak® | Kelp Products International | Природні цитокініни, ауксини, бетаїни (Ascophyllum nodosum) | 2–3 л/га | Кущення, прапорцевий лист | +8–20 % врожай, +10–18 % маса 1000 зерен, сильний антистрес | ↑ sink strength зерна → зерно сильніше “витягує” асимілянти; ↑ транспорт сахарози через SUT/SWEET; затримка старіння листа → фотосинтез триваліший |
| 2 | Cytokin® | Stoller | Synthetic cytokinin, kinetin-подібні речовини | 0.25–0.5 л/га | Кущення–прапорцевий лист | +10–18 % | ↑ sink strength, покращує транспорт сахарози, затримка сенесценції листя |
| 3 | Stimulate® | Stoller | Kinetin, GA, IBA | 0.3–0.5 л/га | Вегетаційний період | +10–20 % | ↑ sink strength, підтримка транспорту асимілянтів, тривала фотосинтетична активність листя |
| 4 | Algaren S90 | BioAtlantis | Ascophyllum nodosum | 2–4 л/га | Кущення–фаза прапорцевого листа | +8–18 % | ↑ sink strength, стимуляція транспорту сахарози, антистрес ефект |
| 5 | Goëmar BM86 | Goëmar | Природні цитокініни | 2–3 л/га | Кущення–прапорцевий лист | +5–15 % | ↑ sink strength, підтримка транспорту, затримка старіння листя |
| 6 | MaxCel® | Valent BioSciences | 6-BAP (чистий 6-бензиламінопурин) | 0.05–0.1 % розчин | Садівництво/зернові | +10–15 % на зернових | ↑ sink strength зерна, активує SUT/SWEET, затримка сенесценції листя |
Ключовий принцип дії цитокінінових препаратів полягає в посиленні механізму SOURCE → SINK: вони підвищують силу акцепторних органів (sink strength), активізують транспорт сахарози та затримують старіння листя, що забезпечує більший накопичувальний потенціал зерна, крохмалю та білка. У результаті це реально приводить до +8–25 % приросту врожайності пшениці. Реальний приріст урожаю в різних культурах становить: пшениця – +8–25 %, кукурудза – +10–30 %, соняшник – +10–22 %.
Таким чином, наведена інформація демонструє фундаментальний механізм дії цитокінінів і може слугувати базою для розробки проривних антистресових технологій у сільському господарстві.
Ауксини, абсцизова кислота, гібереліни у гормональному контролі SOURCE–SINK.
Ауксини — ключові регулятори транспорту асимілятів і розвитку органів-акцепторів. Вони модулюють потік сахарози через флоему, сприяючи ефективному завантаженню SINK-органів. Ауксини підвищують активність SUT і SWEET, стимулюють ріст меристем, розвиток пагонів і кореневої системи, що забезпечує збільшення сили sink. Практично ауксини застосовують для стимуляції росту пагонів, підвищення урожайності і покращення розвитку кореневих систем. Надлишок ауксинів може призводити до нерівномірного росту і пригнічення формування плодів.
Абсцизова кислота (ABA) діє як негативний регулятор SINK при стресових умовах. Вона пригнічує активність транспортерів сахарози і cell wall invertase, знижує швидкість наливу зерна та обмежує розподіл асимілятів. ABA активується при посусі, високих температурах і дефіциті води, переводячи рослину у режим енергозбереження. Практично використання ABA пов’язане з регуляцією стресових відповідей: контроль над її рівнем дозволяє пом’якшити пригнічення SINK і зменшити втрати врожаю.
Гібереліни — позитивні регулятори росту зерна і репродуктивних органів. Вони стимулюють поділ клітин і розтягування тканин, підвищують активність фотосинтезу та метаболізм сахарози, а також взаємодіють із цитокінінами для збільшення sink strength. Практично гібереліни застосовують для стимуляції росту зерна, підвищення його маси та покращення наливу. Вони особливо ефективні у комбінації з цитокінінами та ауксинами для максимізації продуктивності.
Таким чином, гормональний контроль SOURCE–SINK забезпечує інтеграцію молекулярного транспорту сахарози з фізіологічними процесами росту та розвитку. Цитокініни посилюють силу sink і налив зерна, ауксини координують транспорт і розвиток акцепторних органів, ABA обмежує SINK у стресі, а гібереліни стимулюють ріст зерна. Взаємодія цих фітогормонів формує оптимальний баланс SOURCE–SINK, що визначає врожайність і стресостійкість рослин.
Висновок.
Найважливішим висновком для практичного управління рослиною в умовах стресу є розуміння того, що стрес — це передусім енергетична криза. Будь-який несприятливий фактор, чи то посуха, високі або низькі температури, засолення ґрунту чи дія гербіцидів, призводить до порушення фотосинтезу, пригнічення метаболізму та зниження синтезу АТФ, що робить рослину вразливою та обмежує її ріст і продуктивність. Тому головне завдання полягає у двох взаємозалежних підходах: або захистити фотосинтез, зберігаючи енергетичний потенціал клітини і мінімізуючи втрати через оксидативне пошкодження, або надати рослині готову енергію у вигляді метаболічних компонентів, таких як амінокислоти, фосфор, магній і інші елементи, що стимулюють синтез АТФ і підтримують життєво важливі процеси. Дотримання цього принципу дозволяє ефективно управляти фізіологічним станом рослин, зберегти їх енергетичний баланс, забезпечити відновлення після стресу та створити основу для стабільного росту і високої продуктивності культур навіть за несприятливих умов вирощування.
Стресостійкість і продуктивність рослин визначаються здатністю клітини підтримувати баланс між процесами росту і виживання, який регулюється фундаментальними енергетичними сенсорними системами TOR (Target of Rapamycin) і SnRK1 (SNF1-related kinase 1). Ці сигнальні шляхи функціонують як центральний метаболічний перемикач: при достатньому енергетичному забезпеченні активується TOR, що стимулює синтез білків, клітинний поділ і формування репродуктивних органів, тоді як при дефіциті енергії активується SnRK1, який пригнічує ріст і переводить клітину у режим збереження ресурсів.
Ключовим фактором, що визначає активність цих систем на рівні цілого організму, є ефективність SOURCE–SINK взаємодії, яка забезпечує транспорт і використання асимілятів. Саме безперервне надходження сахарози з фотосинтезуючих SOURCE-органів до акцепторних SINK-органів підтримує високий енергетичний статус клітин і створює умови для активації TOR-залежних процесів росту і формування врожаю. У протилежність цьому, порушення фотосинтезу або транспорту асимілятів призводить до енергетичного дефіциту, активації SnRK1 і пригнічення розвитку репродуктивних структур.
Таким чином, первинною причиною втрати врожаю при стресі є не саме пошкодження клітин, а порушення системного транспорту і розподілу енергії, що інтегрується через SOURCE–SINK систему і реалізується на молекулярному рівні через сигнальні шляхи TOR і SnRK1. Саме ця інтегрована система визначає, чи буде рослина продовжувати формування врожаю, або перейде у стан фізіологічного пригнічення.
У цьому контексті антистресові препарати слід розглядати як допоміжний інструмент, який може частково зменшувати пошкодження клітин і підтримувати функціонування фізіологічних процесів, але не як засіб прямого управління фундаментальними механізмами енергетичної регуляції. Їх ефективність залежить насамперед від здатності рослини зберігати фотосинтетичну активність, транспорт асимілятів і функціональну активність акцепторних органів.
Отже, управління врожаєм є, по суті, управлінням потоками енергії і асимілятів у системі SOURCE–SINK, які визначають клітинний енергетичний статус і активність центральних регуляторів росту TOR і SnRK1. Підтримання ефективної роботи цієї інтегрованої системи є основою стресостійкості і ключовою умовою реалізації генетичного потенціалу продуктивності рослин. Саме тому ефективні агрономічні стратегії повинні бути спрямовані насамперед на збереження фотосинтетичного апарату, забезпечення транспорту асимілятів і підтримання активності акцепторних органів, що дозволяє мінімізувати негативний вплив стресу і забезпечити стабільне формування врожаю.
Хаблак Сергій, д.б.н., агроном-експерт, агроаудитор, 066-44-266-08
Читайте нас у Telegram









Sumitomo Chemical подала заявки на реєстрацію нового фунгіциду bifemetstrobin у Японії, США та Бразилії
13.07.2026, 11:27
ЗЗР та насіння

Італійські дослідники підтвердили ефективність дріжджового біопрепарату проти несправжньої борошнистої роси винограду
13.07.2026, 09:07
ЗЗР та насіння

Дослідники виявили перспективний біоагент проти фузаріозного в’янення бананів
12.07.2026, 23:41
ЗЗР та насіння

При купівлі вершкового масла приділяйте увагу інформації на маркуванні
12.07.2026, 10:49
Новини